编者按:
炎症性肠病(IBD)发病率自 1990 年以来,在亚洲、非洲、南美和中东的新兴工业化国家持续升高。然而,目前针对炎症性肠病仍缺乏有效的治疗手段。目前的一些免疫抑制剂疗法可能存在治疗失败的情况,引起的免疫抑制可能造成不良反应。
而近期的研究表明,以参与促进炎症后组织修复的特定 G 蛋白偶联受体(GPCR)为靶点,或可成为治疗肠道炎症疾病的新策略。
今天,我们特别编译了发表在 Trends in Molecular Medicine 杂志上题为“Therapeutic Opportunities for Repair GPCRs during Intestinal Mucosal Wound Healing”的文章。希望该文能够为相关的产业人士和诸位读者带来一些启发和帮助。
肠道炎症发作后,G 蛋白偶联受体(GPCRs)在消退阶段的建立方面至关重要。目前肠道炎症相关疗法的治疗失败率较高并且这些治疗容易引起免疫抑制,但是修复 GPCRs 的疗法却有着很好的治疗潜力,这是因为此疗法能在不引起免疫应答的情况下另辟蹊径找到炎症消退方法。
肠道炎症是指急性或慢性加剧的免疫反应,它要么是小肠和(或)结肠粘膜壁受损所导致的结果,要么就是其形成原因。
炎症性肠粘膜环境由肠上皮细胞(IECs)组成,其与免疫细胞、间充质细胞和微生物群相互作用(Box1),这些细胞共同释放出多种可溶性介质,以保护身体免受病原体入侵,并启动愈合过程。一旦威胁被清除,组织就可以通过被称为消退(resolution)的过程得到修复并回归内稳态(见图 1)。
一直以来,消退都被认为是一种通过促炎分子的稀释而产生的被动过程。而现在,人们发现,这是一个在炎症发作阶段触发的复杂而活跃的过程,受到抗炎和促修复介质严格控制,这些介质主要包括一类称作特异性促炎症消退介质(SPMs)的生物活性脂质,它们来源于 ω3 和 ω6 多不饱和脂肪酸1。
SPMs 可分为消退素(resolvins)、巨噬细胞消炎素(maresins)、保护素(protectins)和脂氧素(lipoxins),以及许多通过 GPCRs(最大的跨膜信号蛋白家族)传递的信号分子。
GPCRs 在进化上具有高度保守性,有七个跨膜片段,并且可以与大量配体结合,以接收和处理胞外信息2。在胃肠道中,GPCRs 可以调节许多过程,包括消化、免疫细胞浸润、痛觉和修复。本文的重点是 GPCRs 在肠粘膜修复期间充当 SPMs 受体的作用。
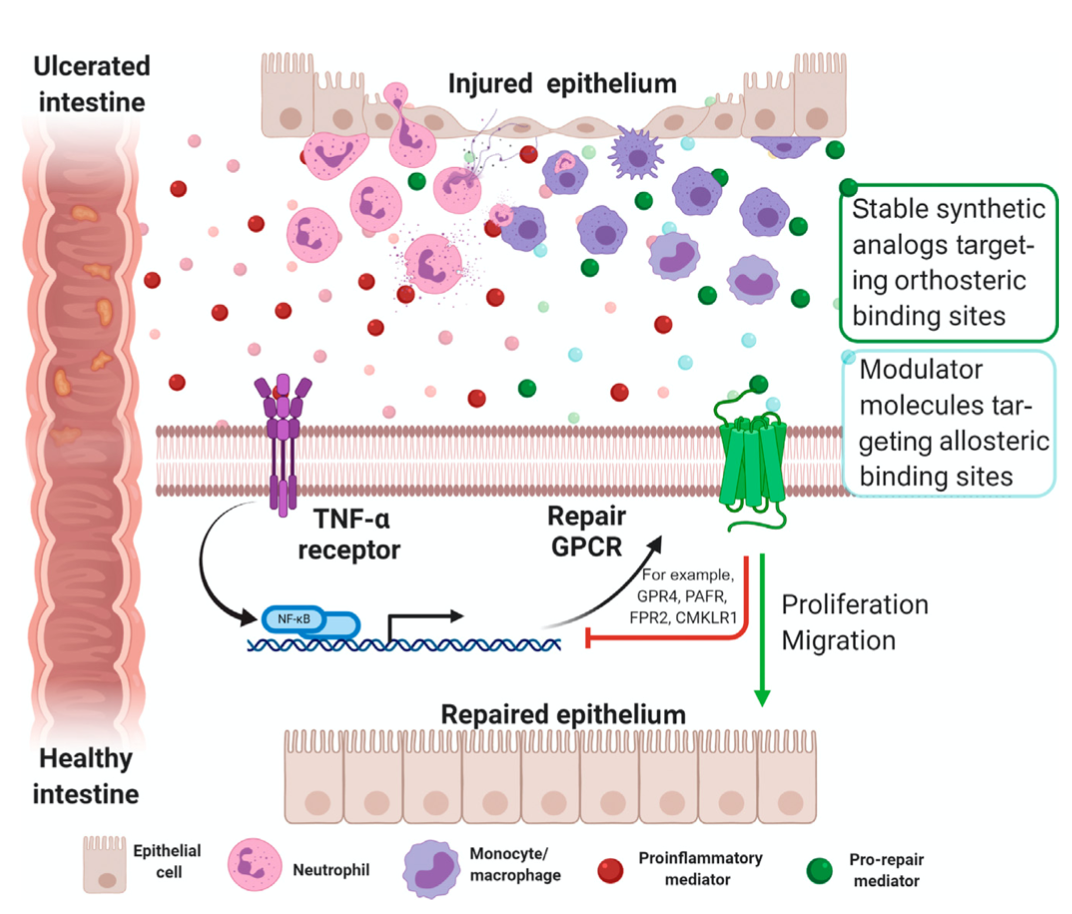
图 1.GPCRs 在肠粘膜修复中的作用。在炎症过程中,上皮膜中的修复 GPCRs 表达量可以通过受损组织中存在的促炎细胞因子(红圆圈)被上调,如 TNF-α。那些具有促修复作用的可溶性介质,如特异性促炎症消退介质(SPMs,绿圆圈),可以通过别构或正构位点结合,以达到修复 GPCRs 的目的,同时也可以促进肠上皮细胞的迁移和增殖,恢复内稳态。那些能够复制 SPM 受体亲和力和促消退作用的稳定的类似合成物,以及通过内源性或合成配体靶向替代结合位点以增强或减少受体激活的调制器分子(浅蓝色圆圈),在治疗方面具有重要意义。缩写词:GPCR,G蛋白偶联受体;SPM,特异性促炎症消退介质;TNF,肿瘤坏死因子。
肠道粘膜伤口愈合是由那些与 IECs、常驻免疫细胞、招募免疫细胞、微生物群和其它固有层细胞(如间充质干细胞 MSCs 和成纤维细胞)相互作用的可溶性介质之间的时空相互作用而协调完成的。
上皮细胞:作为对损伤的反应,与受损粘膜相邻的 IECs 通过集体迁移,来覆盖受损粘膜表面。在这过程中,它们被拉平,失去极性,并经历了细胞骨架的重塑,以实现向前移动达到覆盖伤口的目的。
中性粒细胞:除了介导抗菌宿主防御外,中性粒细胞还可以作为促进粘膜修复的双刃剑。中性粒细胞衍生蛋白酶可以裂解肠上皮连接蛋白,这些裂解产物可以激活旁分泌信号,促进周围 IECs 的增殖。中性粒细胞也产生生长因子,如血管内皮生长因子(VEGF)和 SPMs。
巨噬细胞:在第一波中性粒细胞流入伤口后,巨噬细胞进一步促进宿主防御和修复反应。巨噬细胞通过吞噬中性粒细胞,防止其释放蛋白质,从而抑制组织损伤。据报道,创面相关巨噬细胞会通过分泌肝细胞生长因子(HGF)、TNF-α、IL-1、IL-6、IL-10、IL-12 和 SPMs 来促进宿主防御和伤口修复。
其它免疫细胞:肥大细胞,嗜酸性粒细胞,淋巴样细胞等免疫细胞通过分泌促进修复的生长因子和细胞因子,在组织黏膜修复中发挥重要作用。
固有层基质细胞:MSCs 分泌生长因子和细胞因子,将祖细胞招募到损伤部位,以此介导伤口修复。前列腺素 E2 是 MSCs 分泌的最重要的介质之一,其通过激活 IECs 前体中的 Wnt 信号以维持结肠增殖。肠创面中出现的成纤维细胞也会分泌可溶性介质,如 TNF-α、IL-1β、IL-4 和 VEGF。这些分子可以促进成纤维细胞向创面迁移。创面相关成纤维细胞又可分化为肌成纤维细胞,产生收缩力,促进伤口愈合。
微生物群:肠道菌群能积极调节与黏膜屏障功能相关的蛋白质的表达。无生殖小鼠减少了结肠上皮祖细胞的再生。特定的微生物类型会在肠黏膜创面中富集,并能激活上皮细胞信号通路以促进修复

SPMs 可以激活 GPCRs 并使其保持高度活跃,从而产生强烈的促炎症消退反应,如下调促炎介质(包括IL1β、IL-6、IL-7、IL-8 和肿瘤坏死因子 TNF-α),减少白细胞向炎症活动部位的聚集,并通过浸润单核细胞和巨噬细胞来增强中性粒细胞的凋亡和吞噬作用(细胞吞噬作用)。
SPM GPCR 信号还通过促进 IECs 3的集体迁移和增殖来控制肠上皮黏膜创面修复。
迄今为止,已知的 SPM 的 GPCRs 受体有 FPR2、CMKLR1、BLT1、LGR6、GPR32、GPR18 和 GPR37 4,5。大多数 SPMs 没有明确的受体,同时有超过 100 种 GPCRs 是孤儿受体(这意味着它们的内源性配体尚不清楚),这为以后探索 GPCR/SPM 配对领域打开研究之门,并且可能会转化为治疗方案,以缓解肠道炎症状态6。
SPMs 和 GPCR 促修复下游信号,使得包括 CREB、c-Jun 和 STAT3 在内的不同通路得到激活或抑制。
这些通路推动了大量的抗炎和促修复反应,如增殖、迁移、DNA 修复,以及细菌和中性粒细胞的吞噬作用;而沉默或抑制这些通路则会使功能失调得到修复,同时造成最终的细胞死亡。
伴随修复 GPCR 的激活作用而来的最为重要的修复效果之一是 IEC 迁移的增加。修复 GPCR 信号对于被视为创面相关上皮细胞(WAEs)的 IECs 而言十分重要。 WAEs 经历了去分化过程(上皮向间充质转变),通过改变细胞-细胞连接和细胞骨架,以加速集体迁移和损伤部位覆盖7。
NF-κB 通路被认为是炎症过程的一个标志性特征,并且与肠道炎症性疾病直接相关,比如炎症性肠病(IBD)。肿瘤坏死因子-α(TNF-α)会触发 NF-κB 信号,这是一种与增强促炎基因表达能力有关的典型通路。
有几个 GPCR 修复受体(GPR4、PAFR、FPR2 和 CMKLR1)都含有 NF-κB 结合位点,因此,炎症条件下这些受体的表达会增强。这些 GPCRs 的信号随后抑制了由 TNF-α 触发的 NF-κB 信号,形成一个负反馈回路,从而有助于减少炎症,同时促进消退和修复。
这是一个非常有趣的发现,因为它表明炎症过程受到严格的调控,意味着那些引起初期炎症爆发的相同可溶性介质其实也参与了一部分消退过程的建立。
值得一提的是,有文献8描述了 NF-κB 的 GPCR 激活作用,并且对于特定 GPCRs 下游的 NF-κB 活化或失活特性可能取决于表达受体的细胞类型以及不同介质的存在情况,这些介质会对受体活性产生不同影响。
上述调节级联对肠道炎症的治疗具有重要意义。目前,TNF-α 的抑制抗体被用于治疗 IBD,但这一治疗常常对许多患者没有效果或在一段时间后失效。对于这种效果不佳的现象,一个可能的解释是,TNF-α 的长期阻断使得 NF-κB 信号被完全废止,从而阻止了 TNF-α 对 GPCRs 的诱导修复能力9。

美国和欧盟批准的药物中,大约有三分之一都以 GPCRs 为靶标10,有一些已经进入治疗胃肠炎症性疾病的试验阶段,然而截止目前,依旧尚未获批。不过显而易见的是,无论是在早期炎症阶段,亦或是消退阶段,GPCRs 在粘膜炎症过程中都起着关键作用11。
此外,修复 GPCRs 疗法甚至可能通过促进组织修复,为慢性炎症提供更多治疗可能性。明确了解促修复 GPCRs 的机制将有助于开发治疗药物,能在保证增强修复能力的同时而又不损害免疫反应。
SPMs 具有无限潜力,因为它们作为免疫消退剂,不仅可以抑制炎症,而且也可以激活促修复通路,以此达到增强创面修复能力,而又不损害免疫反应的目的12。
SPMs 对其受体的高亲和力实际上意味着低剂量配体的强效反应。
然而,这些可溶性介质的合成很复杂,并且一旦生产出来,极易失活。因此,开发稳定的可复制其受体亲和力并具有促消解作用的 SPM 类似合成物,深受研究者追捧。
GPCR 配体结合及信号传递受到不同机制的调控,这些机制可以改变分子对其受体的亲和力以及反应强度。我们需要了解的是,不同配体与特定 GPCR 的结合究竟是如何影响生理反应的。
按理说,GPCRs 应该是治疗许多疾病的灵丹妙药,包括肠道炎症,但却很少有合成药物或改性内源性配体能作为这些受体的有效治疗激动剂/拮抗剂。
造成这一问题的原因之一是缺乏那些可以作用于特定 GPCR 亚家族的“活性”(正构)结合位点的高度选择性配体。在密切相关的 GPCRs 中,正构位点往往高度保守,这很好地解释了为什么这些受体是杂乱的,并且难以实现对特定 GPCR 亚型的高选择性。
若要解决这一问题,一种可行的方法就是先识别分子,这些分子要能作为 GPCRs 整个亚家族的激动剂/拮抗剂,参与类似的生理反应。另一种方法是研究调制器分子,以受体的替代(非正构)结合位点(即别构位点)为靶点,并通过内源性或合成配体增强(正调制器)或减少(负调制器)受体激活作用。
一些配体可以使不同的受体构象保持稳定,并选择性地产生一个趋于特定转导通路的“偏向性”信号。
偏向性激动剂在人类治疗学中具有潜在应用价值,因为在设计分子的过程中,可以使其偏向某一有益通路,或者远离某种可能导致脱靶副作用的有害或非理想通路。例如,为了促进肠黏膜修复,最好设计一种可以激活修复 GPCRs 的分子,它偏向于促进 IECs 迁移和增殖的通路,而非持续将免疫细胞聚集到炎症部位。
但是很遗憾,到目前为止,几乎没有与修复 GPCRs 的偏向性信号相关的信息。
修复 GPCRs 有可能成为促进急慢性肠道炎症修复的重要治疗靶点。靶向修复 GPCRs 疗法可以将肠道炎症治疗过程中的治疗方案转变为与经典抗炎药物协同(或替代)的新型修复疗法。这一点很重要,因为 GPCRs 修复调节剂可能会加速创面愈合,而不是像大多数抗炎药物那样只是下调炎症。
参考文献:
1.Quiros, M. and Nusrat, A. (2019) Saving problematic mucosae: SPMs in intestinal mucosal inflammation and repair. Trends Mol. Med. 25, 124–135
2.Wang, W. et al. (2018) New insights into modes of GPCR activation. Trends Pharmacol. Sci. 39, 367–386
3.Quiros, M. et al. (2020) Resolvin E1 is a pro-repair mole- cule that promotes intestinal epithelial wound healing. Proc. Natl. Acad. Sci. U. S. A. 117, 9477–9482
4.Chiang, N. et al. (2019) Maresin 1 activates LGR6 receptor promoting phagocyte immunoresolvent functions. J. Clin. Invest. 129, 5294–5311
5.Cash, J.L. et al. (2014) Resolution of inflammation: targeting GPCRs that interact with lipids and peptides. Drug Discov. Today 19, 1186–1192
6.Gobbetti, T. et al. (2017) Protectin D1n-3 DPA and resolvin D5n-3 DPA are effectors of intestinal protection. Proc. Natl. Acad. Sci. U. S. A. 114, 3963–3968
7.Miyoshi, H. et al. (2017) Prostaglandin E2 promotes intes- tinal repair through an adaptive cellular response of the epithelium. EMBO J. 36, 5–24
8.Fraser, C.C. (2008) G protein-coupled receptor connectivity to NF-kappaB in inflammation and cancer. Int. Rev. Immunol. 27, 320–350
9.Birkl, D. et al. (2019) TNFalpha promotes mucosal wound repair through enhanced platelet activating factor receptor signaling in the epithelium. Mucosal Immunol. 12, 909–918
10.Sriram, K. and Insel, P.A. (2018) G protein-coupled recep- tors as targets for approved drugs: how many targets and how many drugs? Mol. Pharmacol. 93, 251–258
11.Recchiuti, A. et al. (2019) Roles, actions, and therapeutic potential of specialized pro-resolving lipid mediators for the treatment of inflammation in cystic fibrosis. Front. Pharmacol. 10, 252
12. Norris, P.C. and Serhan, C.N. (2018) Metabololipidomic profiling of functional immunoresolvent clusters and eicos- anoids in mammalian tissues. Biochem. Biophys. Res. Commun. 504, 553–561
原文链接:https://www.cell.com/trends/molecular-medicine/fulltext/S1471-4914(20)30216-1
作者|Miguel Quiros
编译|77
审校|617
编辑|笑咲






