
编者按:
食物过敏的患病率日益升高,而过敏的影响远大于人们所经历的不良反应。许多患有食物过敏的孩子不仅面临着潜在的致命性死亡风险,而且还面临着同学欺负的威胁。近年来的研究发现,生命早期微生物组的建立对食物过敏具有重要影响。
今天,我们共同关注食物过敏与微生物,特别编译了发表在 Nature Outlook 杂志上题为“Microbial ambassadors against food allergies”的文章。希望本文能够为相关的产业人士和诸位读者带来一些启发帮助。
当我们摄入第一口固体食物时,我们的消化道中就已经有一个庞大的微生物委员会在列队欢迎了。这些微生物群会将婴儿早期吃下的食物转化为有益的营养素和代谢物,但是它们也有可能会引发一些不健康的免疫反应,最终导致食物过敏。
许多关于免疫系统的描述都是使用了一个简单的比喻,即免疫系统就像一支随时准备识别和攻击外来入侵者(如细菌)的军队。但事实更为复杂,有些自相矛盾的地方是,细菌似乎在训练我们的免疫系统以区分真正的威胁和那些跟我们饮食以及环境相关的无害外来分子。
纽约西奈山医学中心(Mount Sinai Hospital)伊坎医学院的过敏与免疫学家 Supinda Bunyavanich 说:“生命早期的微生物暴露确实影响了我们免疫系统的运行轨迹。”
生活方式和环境因素会在我们生命早期破坏这些肠道生态系统,甚至这种破坏可能在子宫内就已经发生了。在一些婴儿身上,这种破坏导致的微生物失衡,即所谓的菌群紊乱,破坏了他们的消化道功能以及更广泛的免疫健康,创造了导致宿主过敏性疾病的发病条件。
食物过敏真正的流行原因还未完全了解。这在一定程度是由于很多研究都依赖于患者的自我报告。在这种情况下,由抗体介导的过敏反应往往跟非过敏反应(比如食物不耐受)相混淆,后者往往是由消化功能或者代谢功能障碍引起的。
然而,尽管如此,目前出现食物过敏的人数正在逐渐上升:在某些人群中,食物过敏已经影响了多达 10%的儿童。
位于伊利诺的伊州芝加哥大学(The University of Chicago)的免疫学家 Cathryn Nagler 指出,这种食物过敏者持续增加的现象表明,我们不能单纯从食物过敏原与宿主免疫之间的相互作用来理解食物过敏。
她说:“某些环境因素正在参与调节这种反应,这也是促使我们进一步了解微生物群落的原因。”她和她的同事希望通过研究菌群紊乱的原因和结果,来帮助恢复微生物组健康和提供持久缓释的干预措施。

早在 1989 年,伦敦卫生与热带医学院(the London School of Hygiene and Tropical Medicine)的流行病学家 David Strachan 就提出了卫生假说1:在生命早期接触微生物可以对我们的免疫系统健康产生积极影响。
在观测数据的基础上,Strachan 提出,在更小、更清洁的家庭环境中长大的孩子更容易患过敏性疾病。
在随后的数十年内,卫生学假说已经演变成一个以微生物组为中心的模型,在这个模型中,早期暴露于我们家庭以及环境中的微生物对减轻哮喘、花粉热和湿疹等炎症疾病以及食物过敏的风险起着至关重要的作用。
这些疾病在免疫功能紊乱方面有共同的根源,许多患有食物过敏的儿童在长大后会经历其他炎症反应。
澳大利亚基隆的巴温健康中心(Barwon Health)和迪肯大学(Deakin University)的儿科医生 Peter Vuillermin 说:“就现代环境中的免疫失调而言,食物过敏就像是煤矿坑中的金丝雀。”
编者注:煤矿坑中的金丝雀意为危险的预兆。17 世纪时,煤矿工人需要依靠金丝雀来监测煤矿坑中的瓦斯气体,因为金丝雀对瓦斯十分敏感,当空气中出现瓦斯时,金丝雀就会停止歌唱,从而起到预警的作用。
我们在生命早期使用来自母亲、饮食、家庭甚至包括宠物在内的生命体来组装我们自己的肠道微生物组。因此,似乎存在着一个肠道菌群关键的发育窗口期,在这个窗口期内,微生物群得以稳固建立,而这个窗口期也是可能开始出现菌群紊乱的时候。
举个例子,Bunyavanich 和她的同事在 2016 年的一项研究中追踪了牛奶过敏在幼年时的形成基础,她们在 226 名儿童群体中检测了微生物群的多样性2。
她说:“我们发现,3~6 个月左右婴儿的肠道微生物组成对食物过敏的影响最大。”而超过这一时间点的微生物组成差异表现出较少的影响。
但是,其他研究表明,微生物组在生命的最开始几年中会继续经历有意义的重组。Nagler 说:“很多人都在讨论生命开始的前一千天,到了二、三岁的时候,一个人的微生物组就相对稳定了。”
抗生素也与卫生假说有关。它们的广泛使用貌似是导致微生物组紊乱的主要因素。
Nagler 说:“美国的孩子通常在两岁之前就服用过六种抗生素,而其中很多抗生素是针对病毒性耳部感染的,因此无意间对正在发育中的微生物组造成了很多的破坏。”
她和她的同事在 2004 年开展的一项开创性研究中证明了抗生素与食物过敏之间的联系3。她们通过广谱抗生素来杀灭肠道菌群,诱导幼年小鼠对花生过敏。这一实验还揭示了一种免疫细胞受体似乎可以对微生物产生的信号作出反应,阻止过敏的发生。
但是,菌群紊乱的基础甚至可以在出生时就被奠定。
Nagler 指出,婴儿通过产道的过程中所获得的乳杆菌菌种通常是在婴儿肠道中最早建立自身生态位的微生物种群之一。剖腹产所生产的孩子则不会获得这些细菌,而这有可能会影响后续依赖乳杆菌的产物乳酸作为食物来源的细菌定植。
尽管数据有限,但仍有一些证据表明剖腹产与食物过敏易感相关。其中有一项瑞典研究发现,与经阴道分娩的儿童相比,剖腹产的儿童风险增加了 21%4。通过剖腹产生产的儿童更偏向于从医院环境中获取细菌,其中可能包括病原菌。
此外,母乳喂养在建立微生物组方面也起重要作用:在 4~6 个月时过渡到固态食物时,母乳喂养有助于建立起与成年人更相似的微生物组。
马萨诸塞州的波士顿儿童医院(Boston Children's Hospital)的免疫学家 Talal Chatila 解释说:“微生物组发生了整体的变化,梭菌种类增多。”他补充说:“梭菌属的细菌似乎在建立对食物抗原的免疫耐受中起着特别重要的作用。”
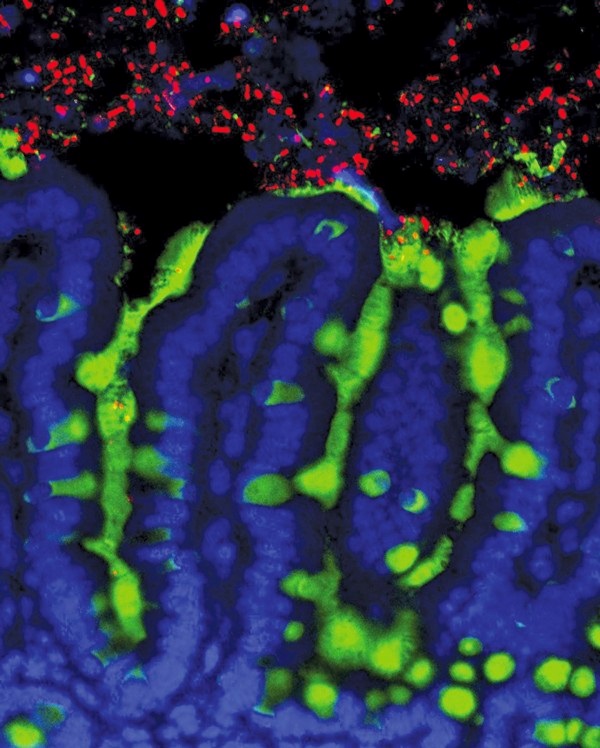
图. 老鼠小肠内的细菌(红色)
对大批儿童的纵向研究使研究人员能监测微生物组结构的变化与食物过敏发生之间的关系。这些研究大多使用了 16S rRNA 测序技术,该技术可以鉴定样品中微生物的 DNA 特征,但通常不够敏感,无法识别具体的菌种和菌株,不过可以揭示导致免疫功能障碍的微生物亚群。
加拿大埃德蒙顿艾伯塔大学(University of Alberta)的流行病学家 Anita Kozyrskyj 与加拿大健康婴儿纵向发展(CHILD)研究进行了广泛的合作,这项研究历时多年,旨在研究 3500 多名儿童的免疫疾病相关因素。
在 2015 年的一项研究中5,她的团队对从 CHILD 队列中的 166 名婴儿收集的 3 个月以及 1 岁的粪便样本进行了 16S rRNA 测序。测序结果揭示了后来被诊断出食物过敏的儿童的微生物群的模式,例如来自肠杆菌科的细菌相对于来自拟杆菌科的细菌更丰富。
Kozyrskyj 说:“我们还发现物种丰富度的降低——微生物组研究中常见的一种测量指标——是危险因素。”
大量的研究表明,梭状芽孢杆菌在调节免疫功能上发挥着重要作用。瑞典于默奥大学儿科免疫学家 Christina West 领导的研究人员在 2019 年的一项研究6中关注了该微生物类别的多个代表菌株,试图将婴儿微生物组的扰动与儿童后期过敏性疾病的诊断联系起来。
West 说:“我们可以看到,从婴儿期到入学年龄这一阶段梭菌纲的粪球菌属一直存在不足,而该纲下的另外一个菌属则只是暂时不足。”她指出,粪球菌倾向于靠近肠道细胞,这表明这些微生物与宿主之间可能存在直接的相互作用。
West 正在寻找更深层次的与微生物相关的过敏因素,他在开展一项名为 NorthPop 的项目来改变导致过敏的因素。这个庞大的纵向人群阵列最终将包括 10000 名来自瑞典 Västerbotten 地区的孕妇及家人。
另外一个纵向研究计划,即澳大利亚的 Barwon 婴儿研究(the Barwon Infant Study)发现了有趣的证据,证据表明孕妇肠道内菌群会影响儿童食物过敏的发生。
在对 316 名妇女及其婴儿的粪便进行分析之后7,研究人员发现,母体肠道中存在的普雷沃氏菌与婴儿到达 1 岁时不出现食物过敏密切相关。
该研究的作者 Vuillermin 补充说:“这种相关性似乎不是由于婴儿在出生后是否携带普雷沃氏菌所引起的。”尽管 Vuillermin 和他的同事仍在努力阐明微生物影响的本质,但这一结果表明普雷沃氏菌可能会通过分泌可穿过胎盘的有益代谢产物来影响胎儿的免疫发育。
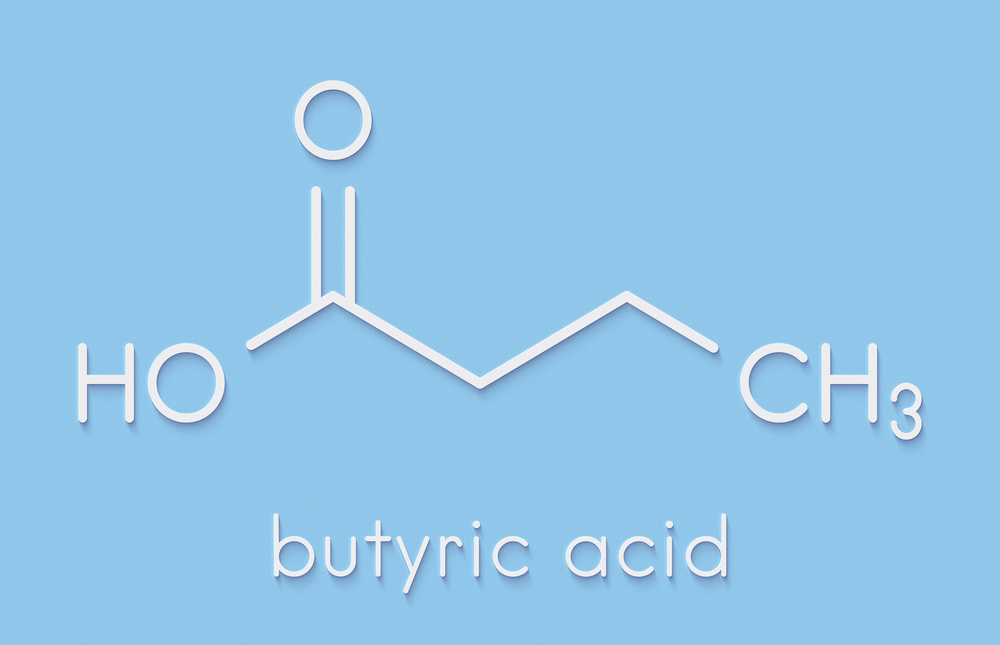
许多研究小组正在努力鉴定肠道微生物与其宿主之间传递的分子信息,以了解它们是如何影响过敏性疾病的。
这些信号中的许多似乎与细菌在代谢肠道食物时产生的化合物紧密相关。特别有趣的是短链脂肪酸(SCFAs),这是一种细菌通过消化膳食纤维产生的分子。就其有益效果而言,丁酸盐是研究最深入的短链脂肪酸,但研究人员还确定了该化学家族其他成员的作用,包括乙酸盐和甲酸盐。
短链脂肪酸和其他细菌代谢产物被认为可以通过不同但彼此相关的机制来影响过敏性疾病发生的风险。第一个机制是它们会在训练免疫系统识别食物中的过敏原(如花生和鸡蛋)时起作用,帮助免疫系统认识到这些过敏原是安全的、不值得产生过激的炎症反应。
Chatila 说:“调节性 T 细胞(Treg)能够对肠内容物,包括食物和细菌产生免疫耐受性。而这些细胞非常依赖于细菌来源的信号。”
菌群紊乱的肠道一般被认为无法产生充足的短链脂肪酸和其他可以充当调节性 T 细胞信号的化合物,从而导致对饮食过敏原缺乏免疫耐受性。
另一个机制是从肠内爆发的过敏原导致的。菌群中的扰动会破坏肠壁的结构完整性,从而将本应紧密的肠道屏障变成多孔的屏障。
West 说:“对食物过敏的婴儿和儿童的肠道有可能会出现渗漏,因此会增加抗原和过敏原的转移。”我们的消化道内部通常不受免疫监视,但如果这种肠道屏障发生渗漏,源自食物的蛋白质会从肠道渗出并引发敌对反应。
据 Nagler 称,一些引起儿童过敏的最常见食物,例如花生和牛奶,都含有稳定存在的蛋白质,这些蛋白质很可能会完整地通过受损的肠壁屏障并引发免疫反应。
她和她的同事在用抗生素处理的小鼠血液中检测到了完整的花生蛋白8。她说,有意思的是在接受抗生素治疗的老鼠中没有梭状芽胞杆菌,这也表明这类微生物可以帮助降低食物过敏风险。
通过对无菌小鼠的研究,已经弄清了微生物组与宿主间关系的许多细节。这些动物在无菌条件下饲养,因此缺乏肠道微生物组,但可以在它们体内定植手动挑选的微生物,以研究这些微生物是如何影响肠道和免疫功能的。
Chatila 的研究小组已利用这类动物来识别特定的信号通路,该信号通路可响应微生物信号而特异性触发调节性 T 细胞的活化。
在 2019 年,他的团队就从人类微生物组中分离出梭状芽胞杆菌类的子集,并将其移植到无菌小鼠中,结果发现可以通过这种途径来抑制食物过敏的发生9。Chatila 说:“你可以治愈这些小鼠的食物过敏,或者是预防它们的发生。”

十多年来,研究人员一直在探索是否可以通过移植一种或多种人工选择的微生物菌种以逆转人类肠道不良状态。然而,在使用此类微生物进行的一些临床研究中获得的经验通常令人失望。
尽管被移植的微生物在婴儿中成功定植,但这种细菌往往会在儿童童年后期死亡。Chatila 说:“当这些孩子到达上学的年龄时,肠道菌群没有差异,免疫反应也没有差异。”这表明移植的微生物可能需要更多的支持,才能成功地建立一个成熟的微生物组群落。
一种可能的解决方案是“合生制剂”配方——将具备潜在治疗效果的菌株和它们最喜欢的食物包装在一起,比如母乳中发现的一些复杂糖类。这个想法的根据是为目标微生物在建立稳定种群方面提供优势。
在 2019 年的一项研究中,Nagler 和同事们对梭状芽胞杆菌类的粪厌氧棒杆菌进行了研究,在无菌小鼠模型中该菌可以单独逆转牛奶过敏10。尽管她的团队还没有找到该细菌理想的食物来源,但她正通过其初创公司(位于芝加哥的 ClostraBio)探索该物种用于合生制剂的潜力。
她说:“你可以在相对较短的时间内使它重新定植在肠道。” 粪厌氧棒杆菌对肠道的许多影响都是通过其生产的丁酸盐来介导的,ClostraBio 公司正在探索其联合创始人 Jeffrey Hubbell 开发的控释制剂将丁酸盐直接输送给患者的潜力。
另一种选择是通过粪菌移植(FMT)重新启动肠道系统,利用供体的健康微生物群替换或修复紊乱的肠道菌群。在无菌小鼠身上的研究已经证明了用这种方法恢复健康免疫功能的可行性,并且这种方法已经在临床上用于修复由致病细菌艰难梭菌感染的患者的微生物组损伤。
但是在免疫系统还在发育阶段的幼儿中进行这种手术会引起更多的安全问题。West 说:“有太多的未知。这项手术的缺点是你有可能将潜在的致病菌转移。”
她的团队正在一项临床试验中探索这种方法的可行性,在这项试验中,剖腹产出生的婴儿将用母亲阴道和肠道微生物的混合物进行处理,以防止潜在的过敏性疾病的发生。
开发治疗方法的基本挑战之一是,对不健康状态的多样性仍了解甚少。事实上,一些数据表明,不同食物的过敏途径是不同的。Bunyavanich 说:“我的观点是,根据食物过敏的亚型不同,治疗方法可能也存在差异。”
尽管存在这些差异,但如果研究人员最终能够确定一组核心微生物物种,从而促进稳定的、受免疫保护的肠道环境建立,那么应该有可能设计出广泛的以微生物组为导向的策略来扭转过敏的发作。
鉴于目前可用的干预措施,如口服免疫疗法(儿童连续数年定期接种诱发过敏的食物),取得了一定的成功,这尤其具有吸引力。
Chatila 说:“这些疗法耗时,有副作用,并且你必须针对不同的过敏原复制它们。这也就急需我们开发一种疗法,这种疗法将不仅可以持续地调整一种过敏原的耐受性,并且可以对任何过敏原的耐受性进行调整。”
参考文献:
(滑动下文查看)
1.Strachan, D. P. Br. Med. J. 299, 1259–1260 (1989).
2.Bunyavanich, S. et al. J. Allergy Clin. Immunol. 138, 1122–1130 (2016).
3.Bashir, M. E. H., Louie, S., Shi, H. N. & Nagler-Anderson, C. J. Immunol. 172, 6978–6987 (2004).
4.Mitselou, N. et al. J. Allergy Clin. Immunol. 142, 1510–1514 (2018).
5.Azad, M. B. et al. Clin. Exp. Allergy 45, 632–643 (2015).
6.Sjödin, K. S. et al. Allergy 74, 176–185 (2019).
7.Vuillermin, P. J. et al. Nature Commun. 11, 1452 (2020).
8.Stefka, A. T. et al. Proc. Natl Acad. Sci USA 111, 13145–13150 (2014).
9.Abdel-Gadir, A. et al. Nature Med. 25, 1164–1174 (2019).
10.Feehley, T. et al. Nature Med. 25, 448–453 (2019).
原文链接:https://www.nature.com/articles/d41586-020-02781-9
作者|Michael Eisenstein
编译|Johnson
审核|617
编辑|笑咲






