
编者按:
2013 年,西班牙人 Katerina Tsilingiri 正式提出“后生元(postbiotics)”的概念,此后,不同的研究人员给后生元赋予了不同的名字及定义。直到 2021 年 5 月,国际益生菌和益生元科学会(ISAPP)发布了后生元共识声明,明确了后生元的定义和范围。那么,后生元,也就是我们通俗认为的“死菌”,究竟是如何发挥作用的呢?
今天,我们共同关注后生元,本文将基于 ISAPP 发表的后生元共识声明文章,对后生元的作用机制进行解读。希望本文能够为相关的产业人士和诸位读者带来一些启发和帮助。
后生元是指对宿主健康有益的无生命微生物和/或其他成分的制剂。由于后生元“无生命”等特征,使其拥有了巨大的应用前景。因此,尽管与益生菌相比,无论是起步时间,还是研究数量,后生元都远远落后,但是学术界和产业界仍为之着迷,产业界更是将后生元视为继益生菌、益生元和合生制剂之后的第四代微生态产品。
然而,任何成功的微生态产品都离不开充分的科学研究。那么,后生元究竟是如何发挥作用的呢?为何死了的微生物也能有健康益处呢?
实际上,后生元所发挥的健康益处可能是由许多不同的机制所驱动的。在某些情况下,这些机制可能与已知的益生菌作用机制类似。这些机制可以独立或联合发挥作用,主要包括以下 5 种:(1)对微生物组的有益调节;(2)增强上皮屏障功能;(3)免疫反应的调节;(4)调节全身代谢;(5)通过神经系统发出信号。
那么,这些机制具体是怎样影响机体健康的呢?涉及到哪些后生元呢?下面,本文将一一阐述。
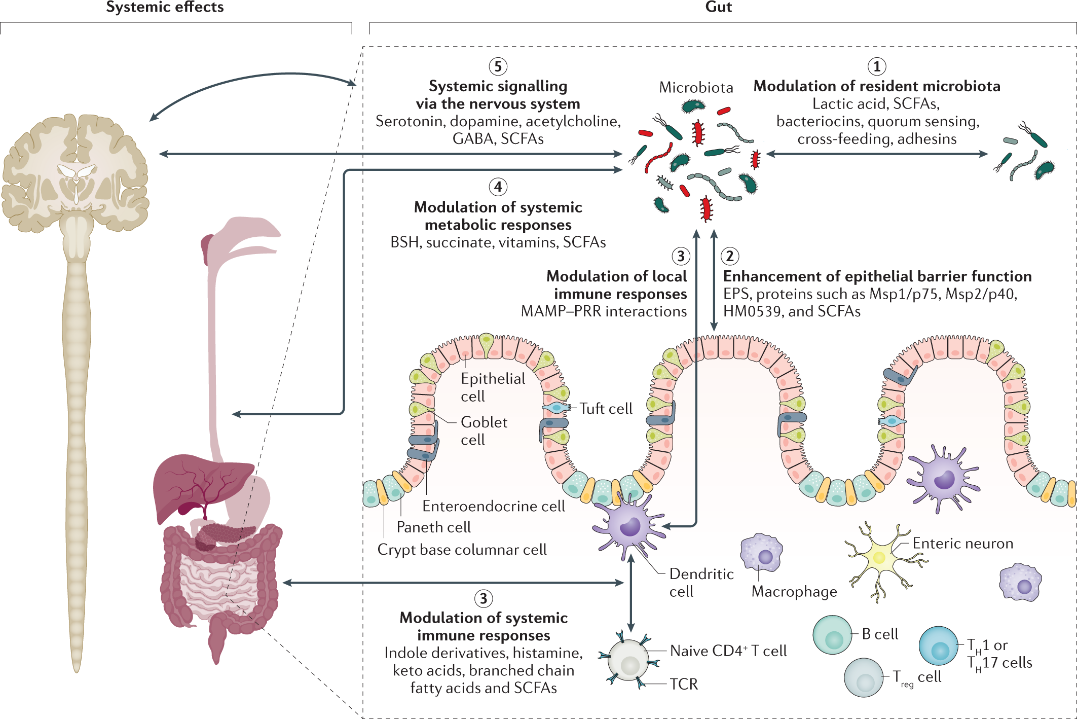
图. 后生元发挥作用的潜在机制及利用该机制的示例。图片来源:Nature Reviews gastroenterology & hematology
虽然后生元对微生物组的影响可能是暂时的,但它们仍然可能具有重要的作用。体内研究表明,后生元中的分子,如乳酸还有细菌素,都具有抗菌活性。
后生元也可以间接地调节微生物组,例如:后生元中可能存在群体感应和群体猝灭分子;肠道中的某些微生物可以利用后生元中携带的乳酸,产生有益的短链脂肪酸(SCFA)和丁酸盐。
如果在处理后,后生元中仍含有保持原样的黏附素(如菌毛和凝集素),那么后生元也可以与常驻微生物竞争黏附位点。
分泌蛋白可介导增强上皮屏障功能,如主要分泌蛋白 Msp1/p75 和 Msp1/p40,或来自益生菌鼠李糖乳杆菌 GG 的蛋白质 HM0539。此外,例如从双歧杆菌中提取的胞外多糖,可以通过尚不明确的信号机制减少炎症,以促进屏障功能。
越来越多的证据表明,特定的双歧杆菌可以激活信号通路,如 MAPK 和 AKT 信号通路,通过自噬和钙信号通路,来促进紧密连接功能。
后生元中的 SCFA 如果达到足够的水平,也有可能会改变上皮屏障功能,并抵御脂多糖诱导的破坏。例如,在 Caco-2 细胞中,丁酸盐可通过组蛋白乙酰化激活脂肪氧合酶,来改变紧密连接的通透性。
局部和全身的免疫调节活动,通常由微生物相关的分子模式与免疫细胞的特定模式识别受体相互作用来介导,从而导致各种细胞因子和免疫调节剂的表达。
当前,通过分子相互作用研究和动物模型验证,已经确定了多种微生物相关分子模式与特定免疫受体的相互作用:
• 脂磷壁酸与 TLR2 或 TLR6 存在相互作用;
• 肽聚糖或其衍生的多肽与 NOD2 存在相互作用;
• 菌毛和纤毛可以调节 TLR2 信号;
• 鞭毛主要与 TLR5 存在相互作用;
• CpG-DNA 与 TLR9 存在相互作用;
• 革兰氏阴性菌(如大肠杆菌 Nissle)衍生的某些后生元脂多糖,主要与 TLR4 相互作用,有时与 TLR2 相互作用;
• 酵母(如酿酒酵母)中的 β-葡聚糖与 TLR2 和凝集素免疫受体存在相互作用;
• 脂蛋白主要与 TLR2 存在相互作用。
如果不被灭活过程破坏或改变,这些与微生物相关的分子模式也可能存在于后生元中。有研究表明,一些来自革兰氏阳性菌和革兰氏阴性菌的裂解物,可以与 TLR4 和 TLR2 相互作用。
此外,据报道,乳酸等代谢物可通过肠道 CX3CR1+细胞中 GPR31 依赖的树突突起,介导免疫反应。同样地,罗伊乳杆菌产生的色氨酸衍生物——吲哚,能激活小鼠肠道中的 CD4+ T 细胞的芳香烃受体,诱导其分化为 CD4+CD8αα+双阳性上皮内淋巴细胞。
后生元中可能还存在着其他免疫调节微生物代谢物,这些代谢物包括:组胺、支链脂肪酸和 SCFA(已被证明会影响许多免疫反应,包括抑制 NF-κB)。
后生元对全身代谢反应的影响,可直接由灭活微生物内部和表面的代谢物或酶介导。
一个例子是胆盐水解酶(BSH)。这种微生物酶负责胆汁酸的解离,从而使微生物进行进一步的生物转化,并使系统循环的胆汁酸池多样化。
BSH 主要在微生物的细胞质中表达,但也观察到了胞外形式,并且其活性已在益生菌——约氏乳杆菌的过滤上清液中得到证实。而且研究表明,BSH 的丧失,使人更容易复发艰难梭菌感染,而通过粪菌移植恢复这种酶的活性,被证明有助于治疗这种感染。
另一个例子是琥珀酸(一种细菌进行碳水化合物发酵的中间产物)。琥珀酸是肠道糖异生的底物,已被发现可以改善小鼠的血糖调控。
其他已知的宿主代谢调节剂,包括微生物源的维生素和 SCFA。丙酸能改善胰岛素敏感性、糖耐量以及脂质代谢;丁酸可以上调抗氧化剂谷胱甘肽,并能对健康人结肠的氧化应激产生有益的影响。
微生物可以产生多种神经活性物质,这些物质可以作用于肠道和中枢神经系统,可能可以调节动物和人类的行为及认知功能。这些物质包括几种神经递质(如血清素、多巴胺、乙酰胆碱和 γ-氨基丁酸),以及能与大脑中表达的受体结合的各种化合物(例如吲哚和胆汁酸)。
微生物酶也可以代谢营养物质,用于宿主神经递质的合成(例如,色氨酸用于血清素的合成,酪氨酸用于多巴胺的合成),从而降低营养物质的生物利用率。
此外,微生物代谢物也可能可以发挥作用,如 SCFA。SCFA 可以刺激肠嗜铬细胞产生 5-羟色胺;在人体干预研究中,SCFA 能够通过刺激厌食激素的释放,来改善进食行为;小鼠试验表明,肠道中产生的乙酸可以进入大脑,通过中枢代谢机制调节食欲。但是经过处理的后生元中含有的 SCFA 是否还具有上述益处,还需要进一步的研究。
细菌合成的维生素,如B族维生素(核黄素、叶酸和钴胺素)也存在于益生菌产品中,因此后生元也可能还保留着这些维生素。维生素 B 对中枢神经系统功能有重要的有益作用。然而,这些神经活性分子,到底有多少能保留在后生元中,目前还没有详尽的报道。
随着学术界和产业界对后生元关注度的日益增加,无疑,我们将会越来越了解后生元及其发挥作用的机制。而这些关于后生元机制的研究将会进一步推动后生元的应用,助力后生元成为更好用、更安全的益生功能解决方案。后生元时代已经悄然来临,你准备好了吗?
参考文献:
Salminen, S., Collado, M.C., Endo, A. et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat Rev Gastroenterol Hepatol 18, 649–667 (2021).
作者|Jessica、617
审校|617
编辑|咲






