

大家好,我是海南大学食品科学与工程学院张家超。非常高兴能在《肠·道》演讲,和大家分享一些我们课题组关于益生菌在肠道内适应性进化的研究进展。
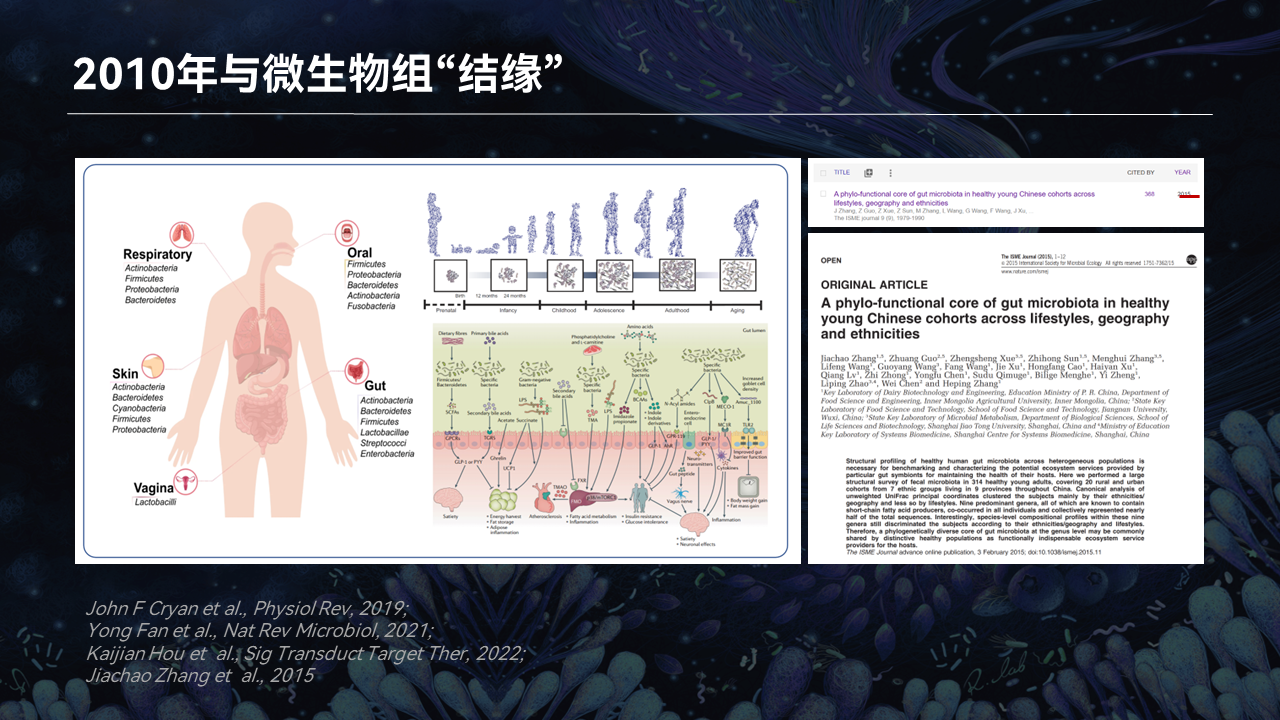
我本人应该算是国内较早一批接触肠道微生物组研究的科研人员。
2010年的时候,西方国家相继在高水平的杂志报道了人类微生物组组成和功能基因的框架,但中国人群肠道微生物组研究仍处于起步阶段。
当时我的导师张和平老师联合江南大学的陈卫老师,还有上海交通大学的赵立平老师,率先开启了中国人群肠道微生物组谱征解析。这个项目历时5年,最终在2015年的时候发表在ISME Journal,我也是这个项目的主要参与者。
现在回想起来,正是在这三位大先生的指引和带领下,我才和微生物组研究结下不解之缘。

我们知道肠道菌群是人体内最复杂的微生态系统。最初大家研究的焦点主要聚集在微生物多样性的层面,也就是“Who is there?”的问题,而后发现肠道微生物组由数千种不同的微生物组成,包括细菌、真菌、病毒。
随着研究的不断推进,研究人员开始探索这些微生物的功能和代谢,也就是“What are they doing?”的问题。大量的研究发现,它们不仅有助于食物的消化、吸收、利用,还有免疫系统的调节,同时还和多种疾病的发生和发展密切相关。然而,肠道内微生物的进化研究仍相对较少。

肠道内微生物之间,它不总是和谐共处的,存在着大量的菌株间对有限资源、生态位的竞争。在竞争的过程中,这些菌株就不可避免地会发生适应性的进化。
深入探索肠道内微生物的适应性进化有着多重的意义。
首先,可以了解生命如何在不同环境中适应和演化,进一步理解微生物与宿主的相互作用。
此外,还有助于我们深入了解微生物演化与宿主疾病的发生发展之间的关联。特别是对于致病菌来说,深入了解其适应性突变对于我们理解致病机制和耐药性机制,这是至关重要的。它也有助于药物的开发和新治疗方法的开发。
既然我们已知微生物在宿主肠道内是会发生适应性进化的,我们课题组的研究的重点又是益生菌,因此这就引发了我们对益生菌是否在肠道内也会发生遗传突变的关注。
这些潜在的遗传突变是否可能对益生菌自身的遗传稳定性、安全性,还有它的功效发挥产生影响呢?这个问题是2015年的时候在我脑海中闪现的。
自2017年起,我们开始专注于探究这个问题。
我们使用了一株植物乳杆菌Lp082作为研究对象。这个菌株是2015年我到海南大学工作以后,从海南省的传统发酵食品中分离获得的。
第一个我们想迫切知道答案的问题就是:外源益生菌到达宿主肠道后,它是不是会发生突变及适应性进化?
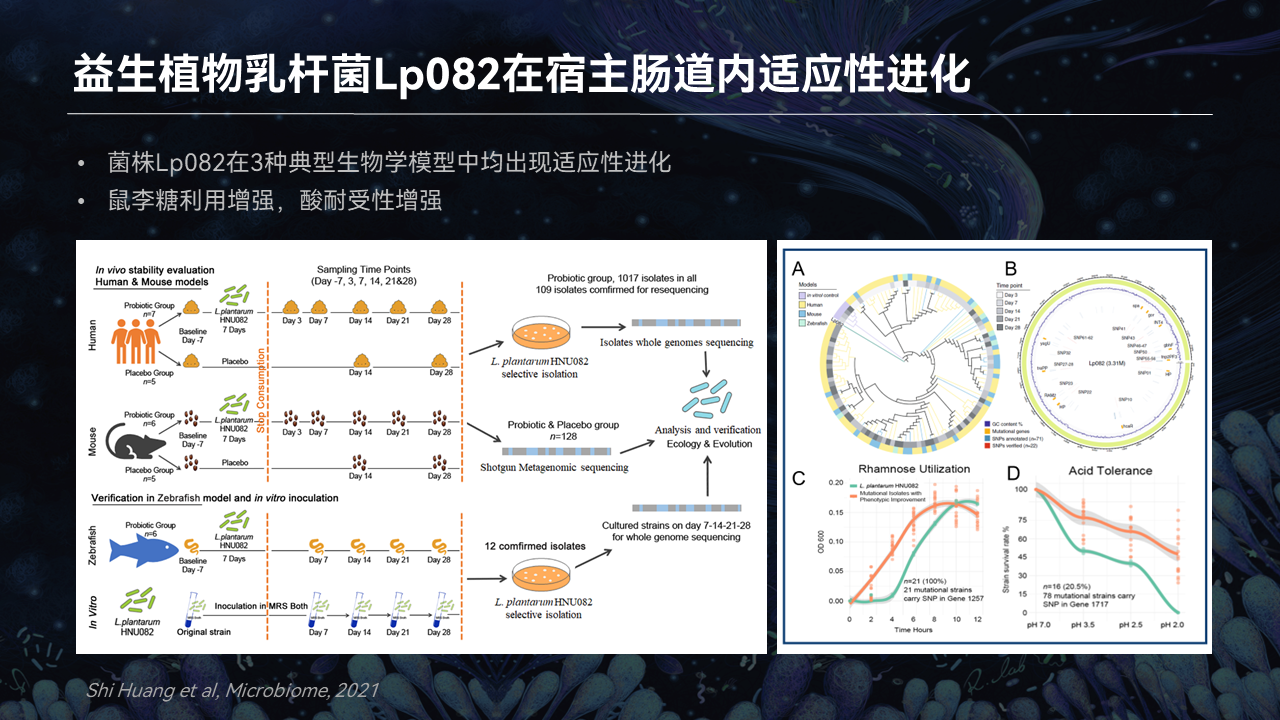
围绕这个问题,我们选择了3种典型的生物学模型,使用益生植物乳杆菌Lp082干预,从肠内容物中分离菌株的突变体,通过对突变位点的注释来探究它在肠道中发生了哪些适应性进化。
研究结果表明,这个菌株在3种典型的生物学模型均经历了适应性进化,并且这些突变涉及到功能层面上,主要表现为鼠李糖利用的增强,同时也展现了更多的酸耐受性。
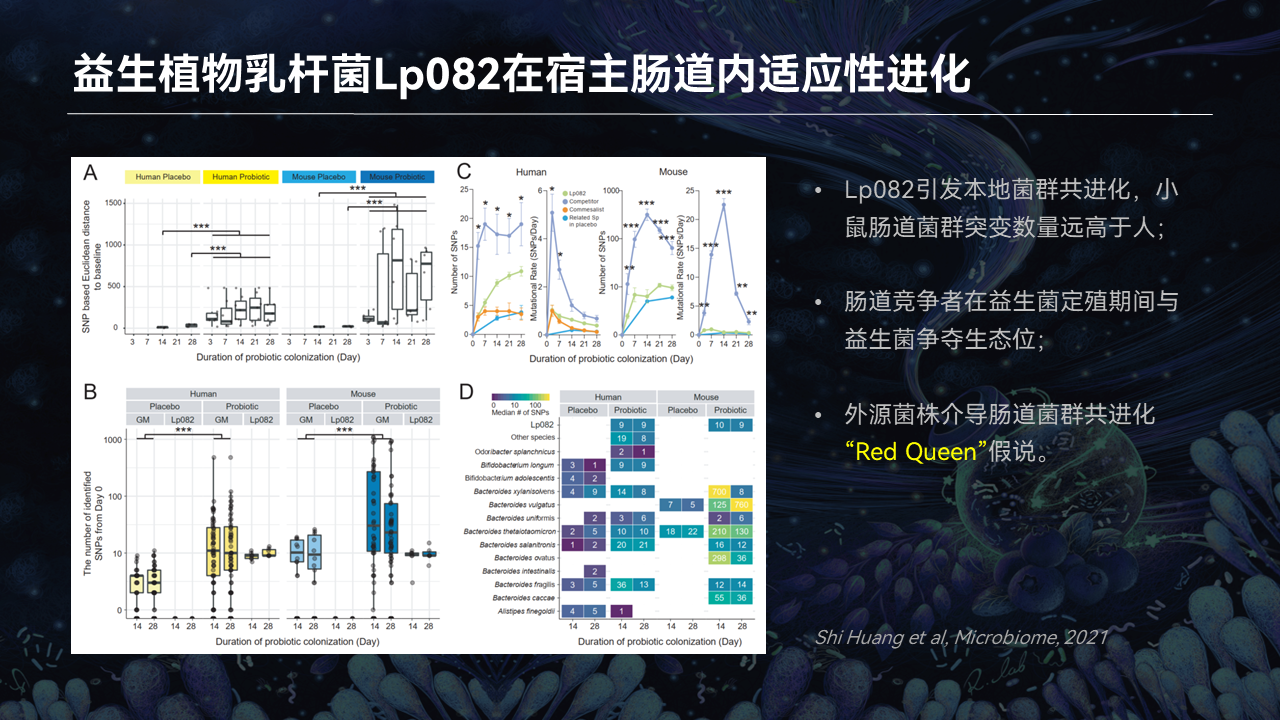
同时我们发现,菌株的摄入在肠道触发了本地菌群的一个共进化现象。
值得注意的是,和人类模型相比,小鼠肠道菌群表现出更高的突变数量。这个可能是和小鼠肠道菌群的稳定性、多样性低于人体肠道菌群相关,所以它更容易被扰动。
而本地菌群大量突变的原因,也可能是在益生菌定植期间肠道中与其竞争生态位的微生物所驱动的。这也表明,生态位和资源的争夺在共进化过程中扮演了重要的角色。
这种现象就和“Red Queen假说”相符合。这种假说提出生物种群必须不断进化,以适应变化的环境和选择压力。本地菌群需要适应益生菌进入肠道后带来的一个竞争压力,而益生菌同样需要不断地进化,让自己适应肠道的环境并在环境中存活下来。
这个是我们在该研究方向的第一个探索性的实验。

上述研究表明,首先植物乳杆菌Lp082在宿主是可以发生适应性进化的,这种进化是由宿主和微生物两方面因素所共同驱动的。而本地菌群在外源益生菌干预的过程中出现了一个共进化的现象。
这个研究就不由得引发了我们更深入的思考。
第一就说,是否所有的益生菌摄入都会引发本地肠道微生物的共进化,也就是说这种现象是不是普遍存在的?
第二个问题就是,肠道内驱动益生菌进化的选择性压力来源于哪里?宿主因素和肠道微生物因素,谁的贡献会更大一些?

为了回答第一个问题,我们就开展了一项荟萃分析,这个分析涵盖了11个益生菌对健康宿主干预的队列研究。
结果表明,首先,所有的益生菌摄入都可以引发本地菌群的一个广泛核苷酸序列的变异。这个发现也证明,益生菌摄入引发本地菌群共进化现象的一个普适性。
同时我们会发现,针对同一个菌株,比如说干酪乳杆菌Zhang,比如说双歧杆菌BV9,同一个菌株对小鼠肠道内引发的这种共进化现象的激烈程度是远高于人群队列的。而这个现象和上述的研究结果相一致,也进一步支持了我们的发现。
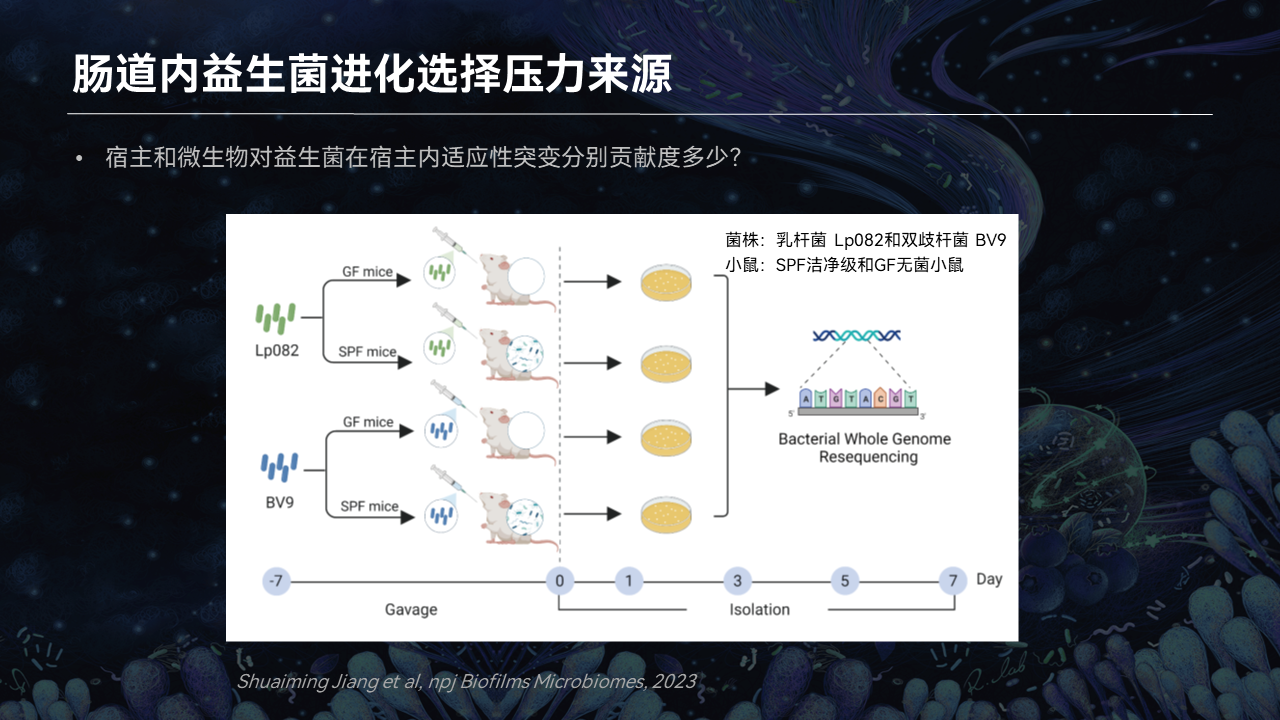
针对第二个问题,我们开展了以下的研究。
为了增加研究的客观性,我们同时选用了植物乳杆菌Lp082、双歧杆菌BV9两种益生菌,分别灌喂给SPF洁净级小鼠和GF级无菌小鼠。通过对分离株突变位点的注释,来探究宿主和肠道菌群分别对益生菌适应性进化所做的贡献。
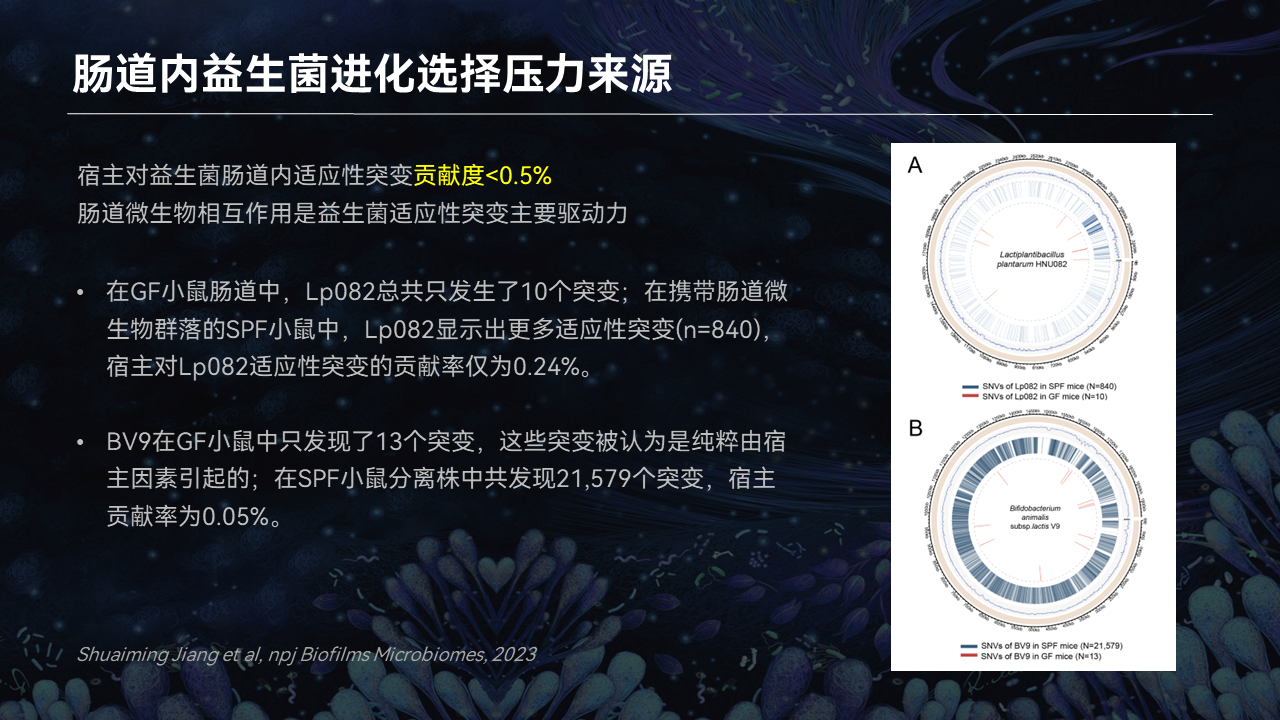
我们发现,宿主因素对益生菌肠道内适应性进化的贡献率仅小于0.5%。所以,它的贡献程度是非常的微弱的。
无论是对于乳杆菌也好,对于双歧杆菌也好,大量的突变引入都源于宿主的肠道菌群所带来的选择压力。
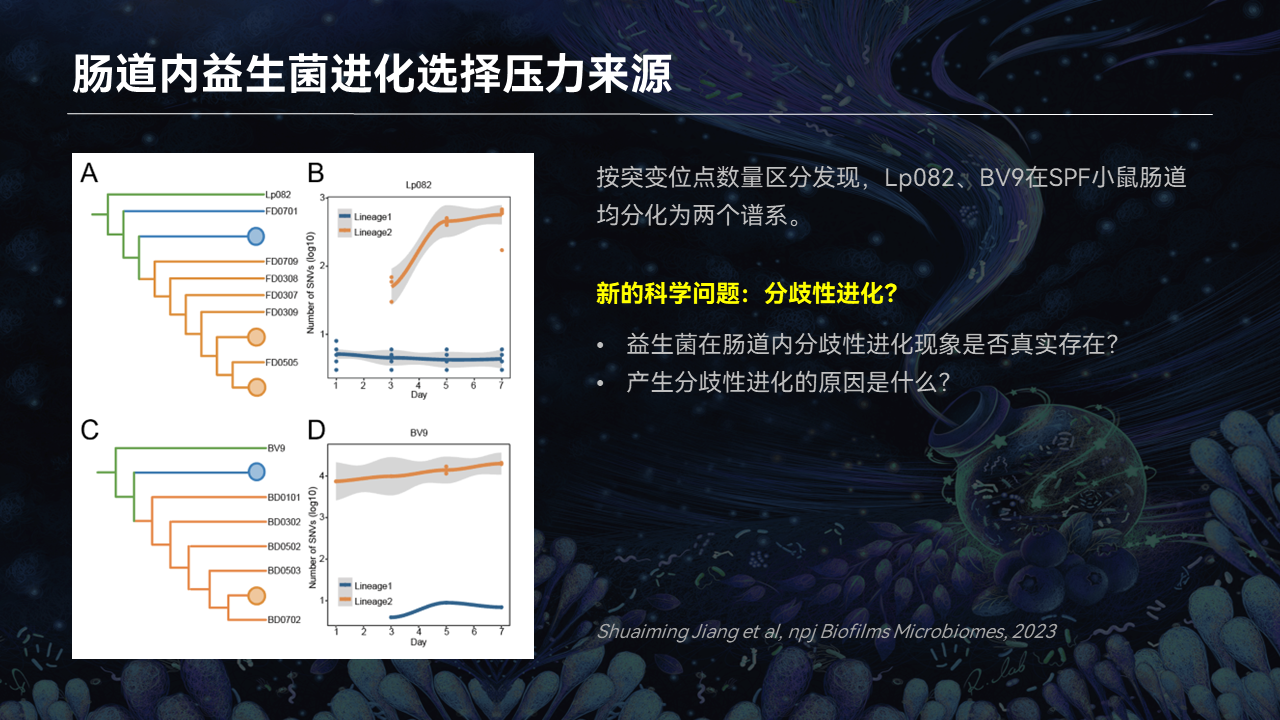
对上述研究中分离到的突变株的突变位点进行数量比较时,我们惊喜地发现,两个突变株在肠道都分化成了两个不同的进化的谱系。这个结果暗示什么?它暗示着益生菌在宿主肠道内采用了一个分歧性进化的策略。
这个观察就引发了一个新的科学问题:益生菌在肠道内分歧性进化策略是不是真实存在的?如果它是真实存在的,那么产生分歧性进化的根本原因又是什么呢?
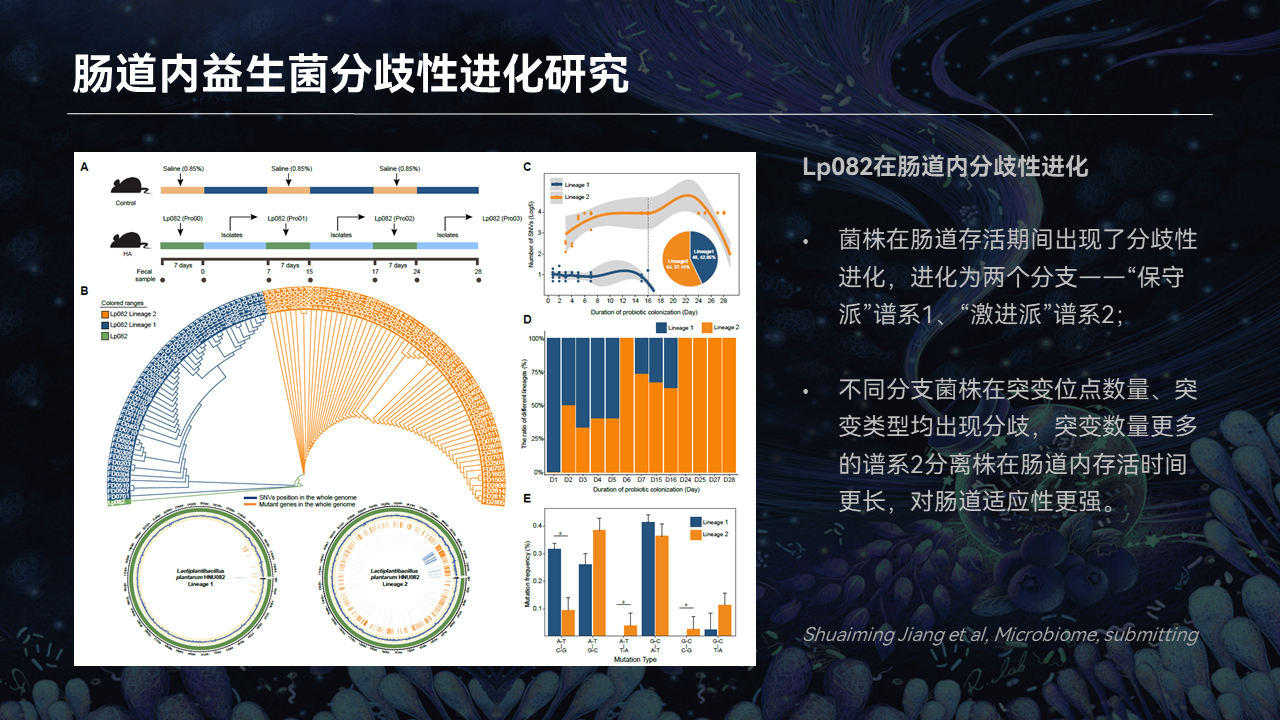
为了回答上述问题,我们依然以植物乳杆菌Lp082为研究对象,通过多轮驯化的实验,来探究肠道益生菌分歧性进化的策略。
具体来说,我们将Lp082菌株首先灌喂给小鼠,而后从粪便分离到它的突变株,再次将菌株灌回给小鼠。经过多轮的驯化,我们对突变株的突变位点来进行注释和分析。

结果发现,菌株在肠道存活期间确实出现了分歧性进化的现象。菌株分为了两个分支,一个是保守派——谱系1,一个是激进派——谱系2。
保守派的突变位点会比较少,激进派的突变位点会比较多,两个分支的菌株在突变位点数量、突变类型上均出现了分歧。而且携带更多突变位点的谱系2,我们发现,它在肠道存活的时间更长,而且对肠道的适应性能也更强。
而后我们对分离株进行系统发育树的构建发现,相比于谱系1,谱系2的分离株与原始的菌株相比,进化距离会更远。而且突变进化图谱展示出,不同分支突变位点的出现时间、存留时间也存在不同。其中lineage 2,也就是谱系2的分离株,会存在多个突变位点得以持续保留的进化现象。
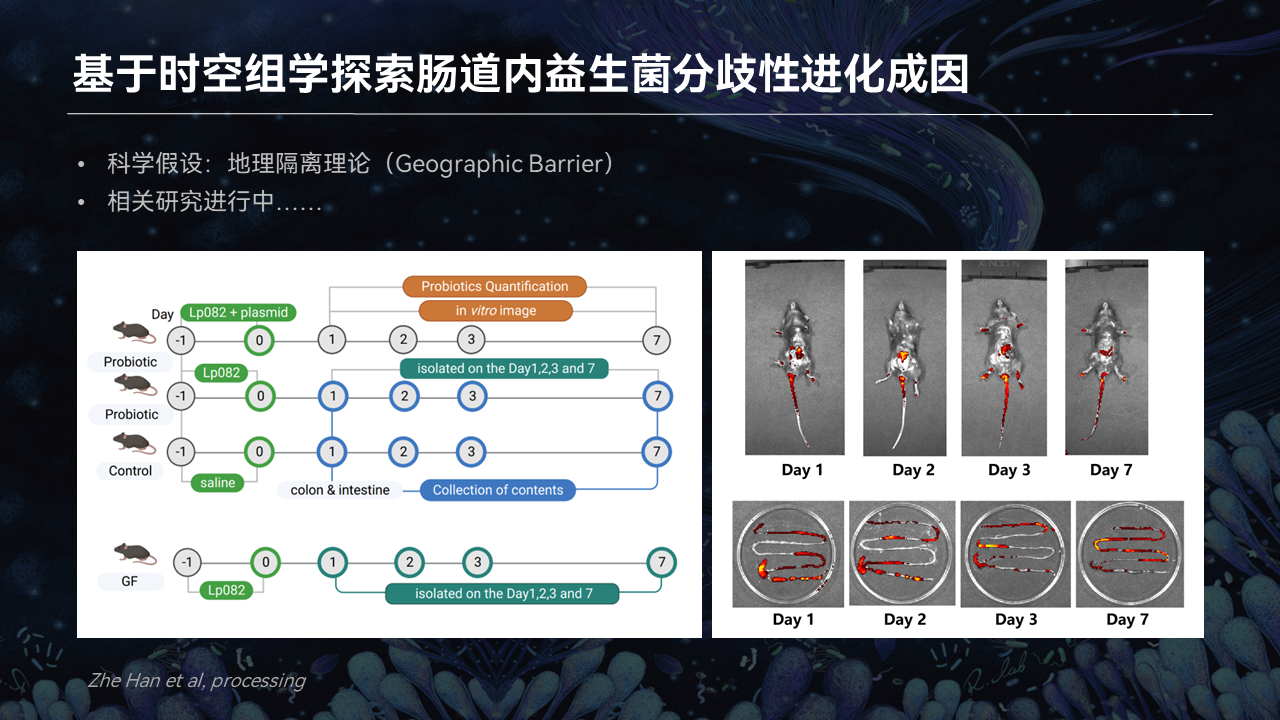
对于造成肠道内益生菌分歧性进化的成因,我们基于时空组学探索了Lp082在进入小鼠肠道不同的肠段后的一个突变情况。
我们初步的科学假设和地理隔绝论是密切相关的。我们认为,益生菌定植在小鼠不同的肠段受到的选择压力不同,是导致它出现分歧性进化的一个根本原因。
我们初步实验的结果也表明,植物乳杆菌进入肠道后主要是在小肠内定植的。而从小肠到大肠分离到的突变株,它的进化速率是否存在显著差异,以及菌株在不同肠段承受选择压力的定量分析,这个研究我们仍在进行中。
通过上述的多项研究,我们逐步地揭开了肠道内益生菌适应性进化的神秘面纱,并且也得到了一些初步的研究结果。
首先,益生菌在肠道是可以发生适应性进化的,主要的进化动力是来源于菌群。同时益生菌在肠道内存活部位不同,而地理隔绝论可能是指导益生菌发生适应性进化的一个根本原因。

所以这些问题就引发了我们进一步的思考:是不是可以基于肠道微生物组来定向驯化益生菌?
围绕这个问题,我们又设计了两个研究。
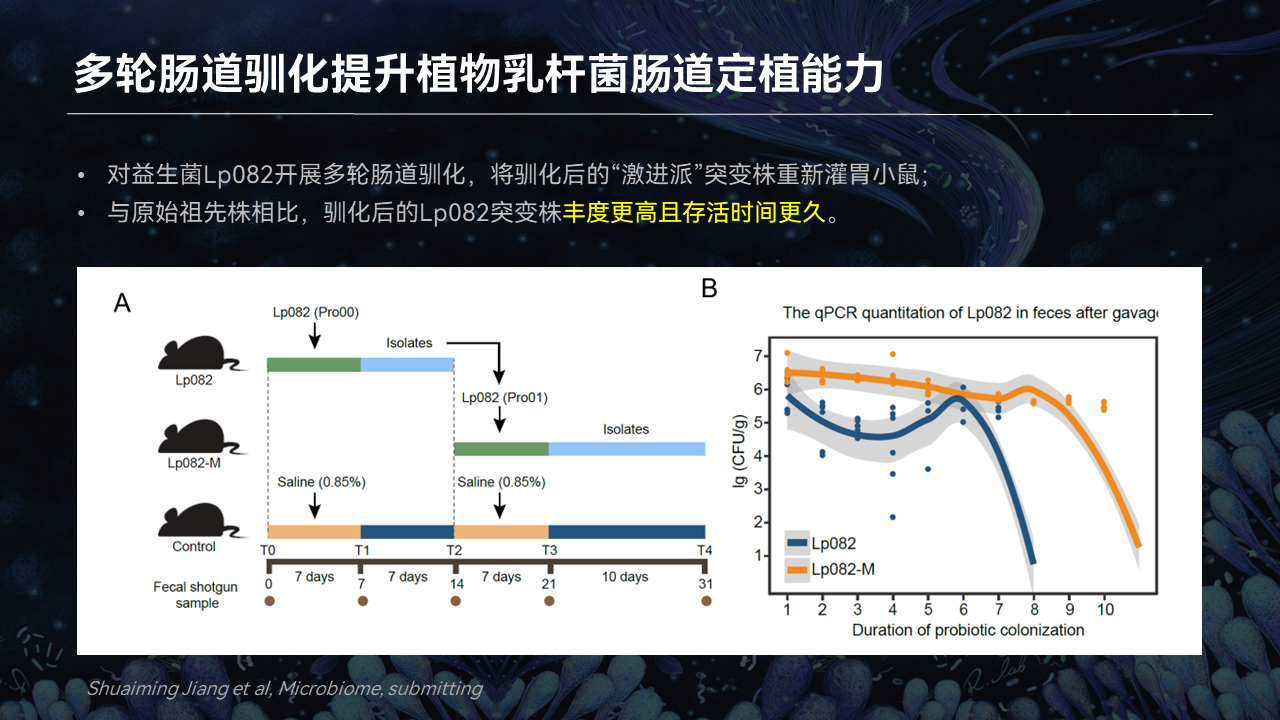
一方面,我们开展多轮肠道驯化,来探究是不是可以提升植物乳杆菌在肠道内的定植能力。
另一方面,通过不同饮食模式干预策略改变肠道的选择压力,我们来看看是不是能够对双歧杆菌的肠道适应性有所提升。
针对第一个思考,结果表明,经过多轮肠道驯化的植物乳杆菌进入小鼠体内,与原始菌株相比,它存活的时间会更久,它存在的丰度也更高。
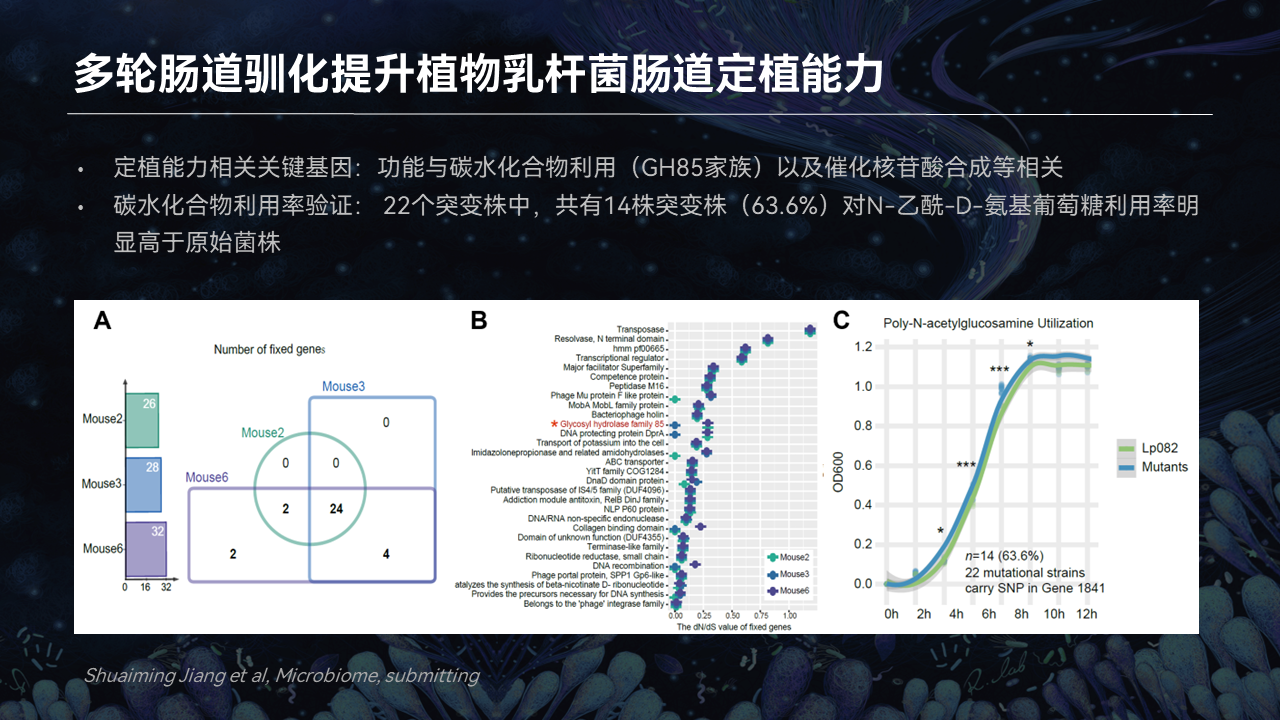
同时这些突变株中与定植能力相关的关键基因功能,主要是与碳水化合物利用、催化核苷酸合成密切相关。
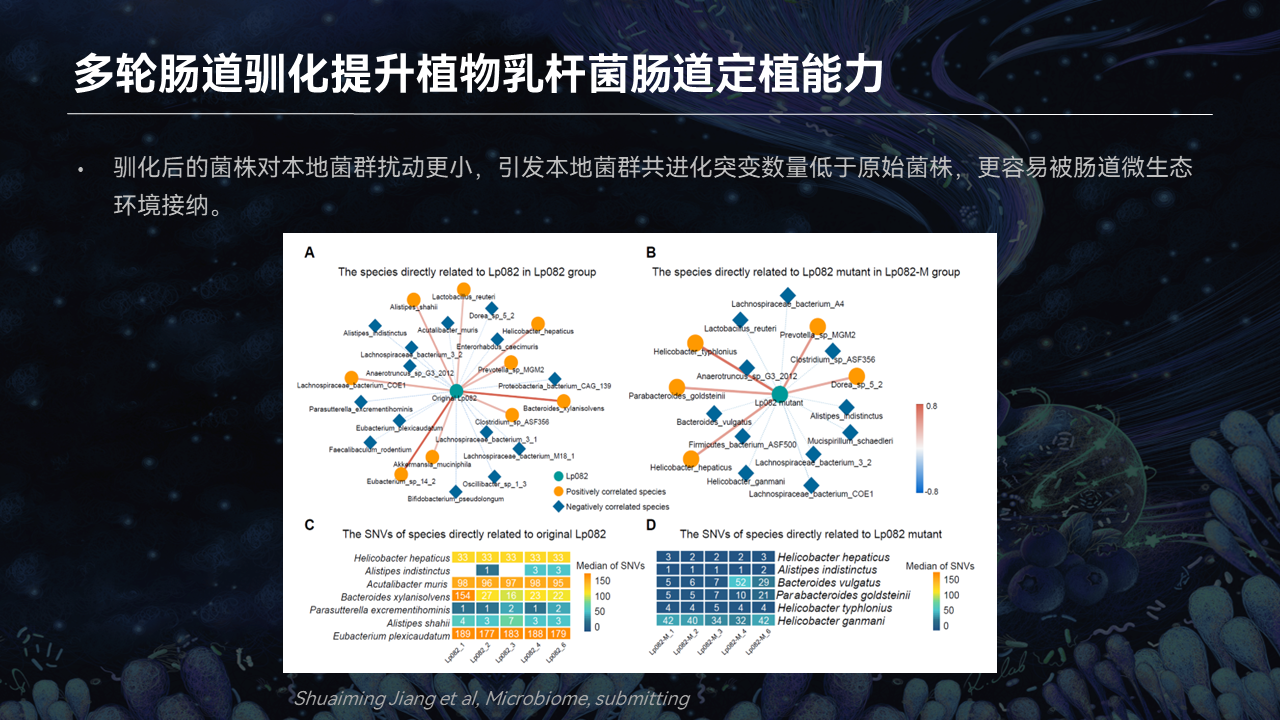
同时我们发现,驯化后的菌株再次进入到肠道后,它引发的菌群突变数量是比原始菌株要远远低的。这意味着驯化后的菌株对本地菌群的扰动会更小,它更容易被原始的肠道微生态环境所接纳。
这是我们基于肠道菌群开展益生菌驯化的第一个研究。

针对第二个思考,我们选用了双歧杆菌BV9来作为实验菌株,将小鼠分为了3个不同的实验组,主要包括了高脂饮食组、高膳食纤维组和正常饮食组。
两周以后在这个基础上,我们将灌喂益生菌的BV9重新从粪便中将它分离出来,然后完成宏基因组测序和单菌株的全基因组测序。
我们这个研究就是想看一看饮食模式所介导的肠道选择压力,是不是能够驱动双歧杆菌产生适应性进化。
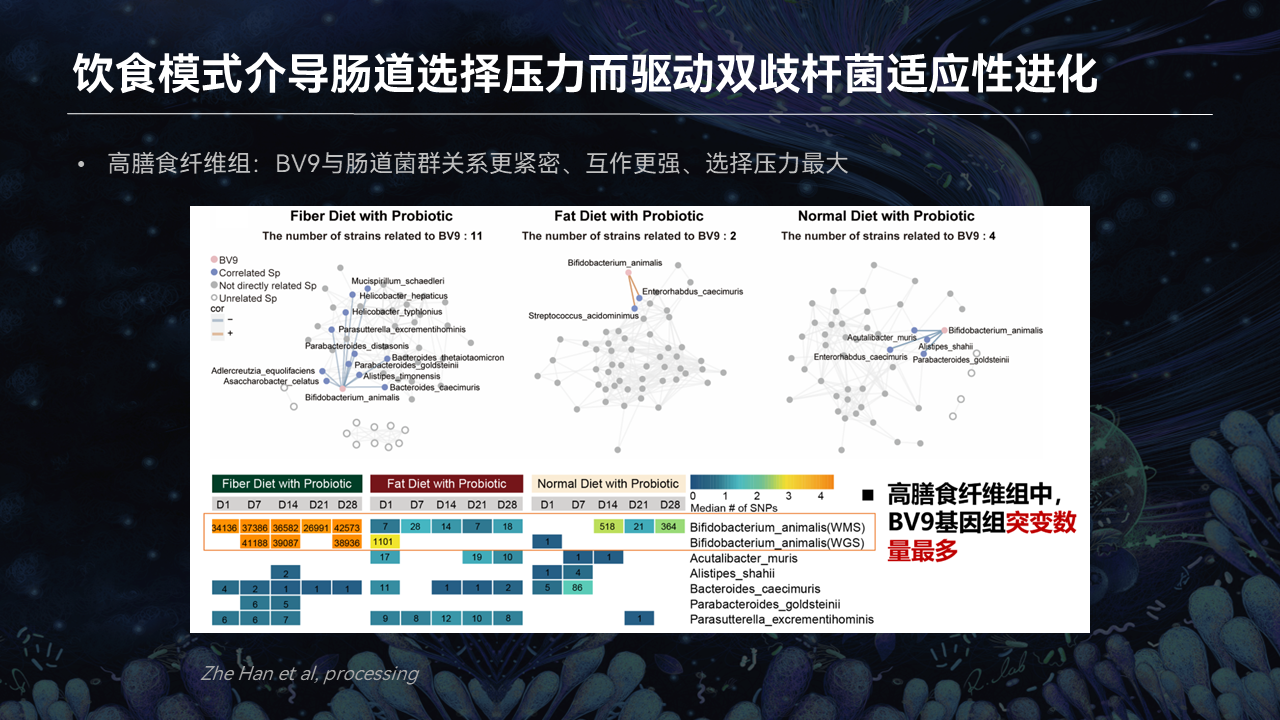
首先我们在建立了3种饮食模式下,益生菌干预后微生物与菌株的一个共生的网络,来探究益生菌和肠道菌群的共进化情况。
有趣的是,我们发现在高膳食纤维组,双歧杆菌BV9和它产生关联的菌株数量最多,益生菌和肠道菌群的关系就更加密切,互作就更强。
此外,在高膳食纤维组,BV9这株外源的益生菌产生的基因组突变数量最多。这也就意味着在高膳食纤维组,BV9受到的肠道选择压力就更大。
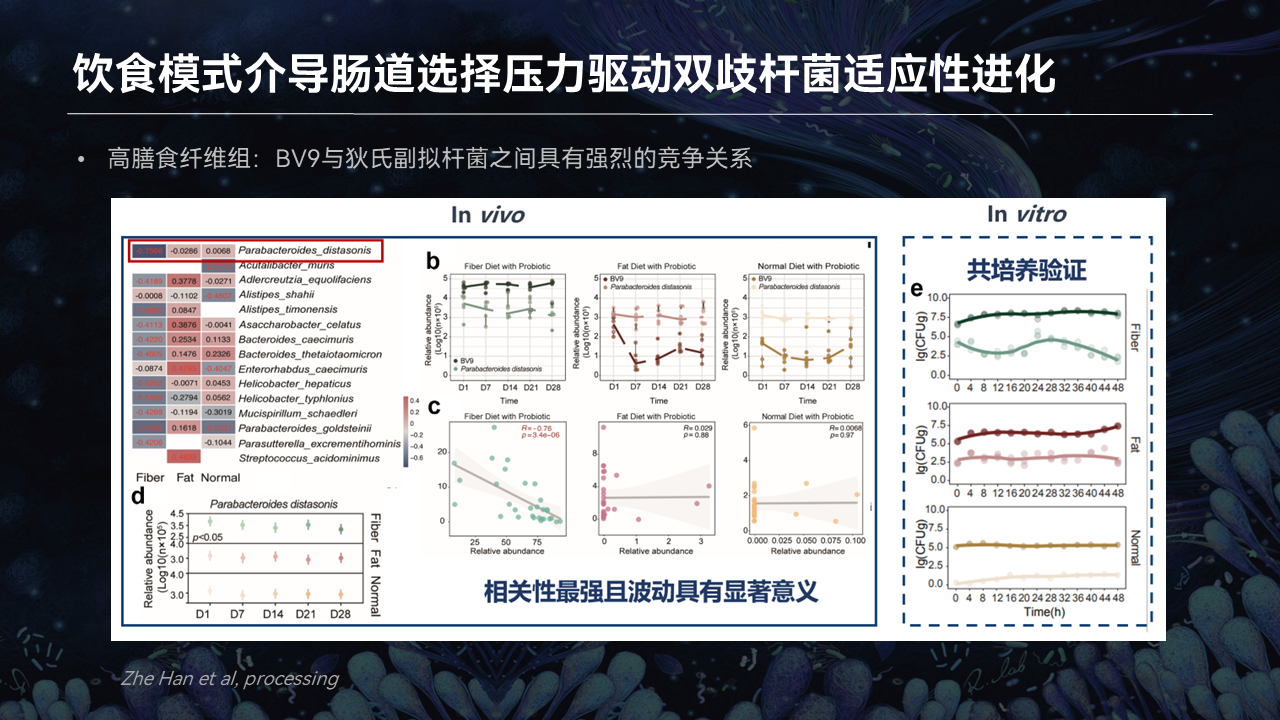
有趣的是,我们发现了一个比较特别的菌种——狄氏副拟杆菌。在高膳食纤维组,狄氏副拟杆菌和BV9之间存在着一个激烈的竞争关系。
首先在高膳食纤维组,饮食塑造的肠道菌群随着时间的推移,狄氏副拟杆菌和BV9在丰度上出现了一个此消彼长的现象。在其他的两种饮食模式中,我们并没有发现这样的现象。
而且在体外共培养的实验,我们也验证了这样的一个现象,同样有这样的一个规律。
那么BV9这个外源益生菌和本土的狄氏副拟杆菌,在生态位上的激烈竞争是不是由竞争性共进化所驱动的呢?
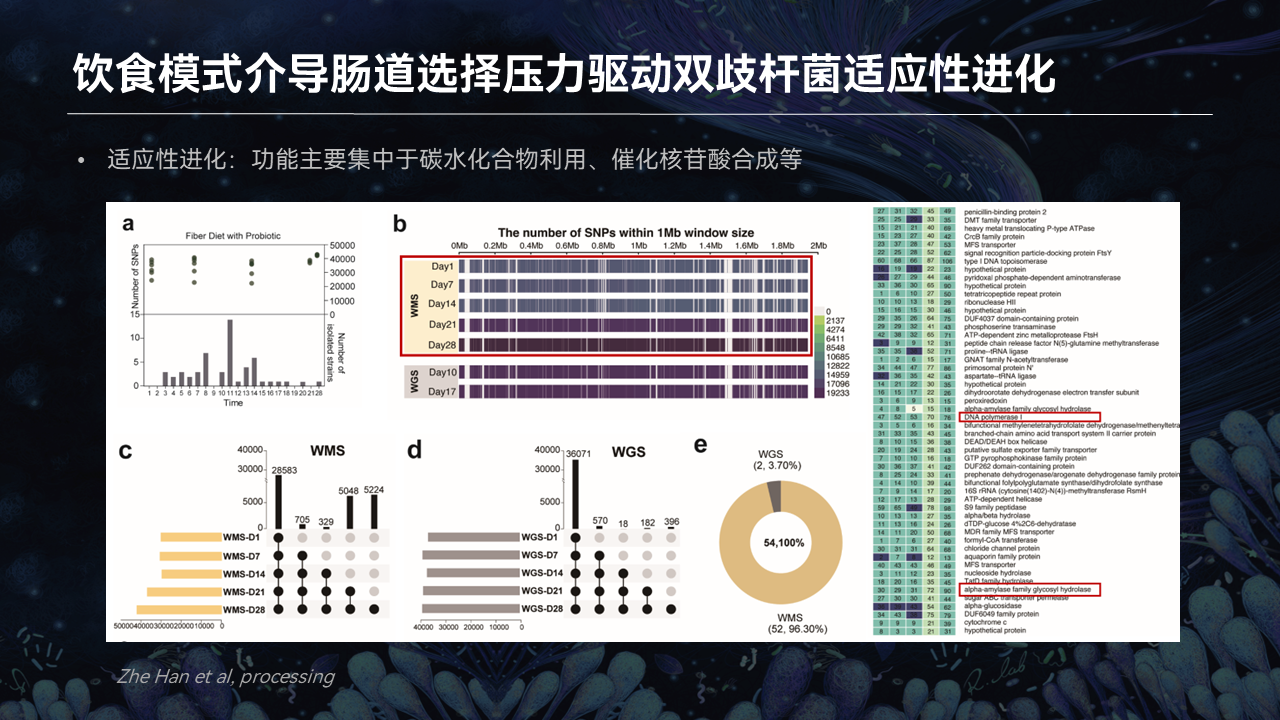
我们就将分析继续下探到了基因组的层面。在整个的实验周期,我们发现,BV9突变位点的数量随着时间的增长而逐渐的增加。
我们还筛选到了一些与定植相关的位点,这些位点随着时间的累积而稳定存在。经过注释,这些位点主要存在于54个基因上,功能主要集中于碳水化合物利用、基因组的复制相关。
而与此同时,我们在狄氏副拟杆菌的基因组等位基因上也注释到了类似的突变。
这就表明,通过高膳食纤维饮食模式,选择性地激发肠道本地菌群与益生菌展开一个竞争性共进化,这种方式能够有效地提升双歧杆菌的肠道适应性。这也证明饮食干预策略驯化益生菌的一个可行性。
以上是我们课题组对益生菌在肠道内适应性进化的一系列探索。

回顾我们课题组整个的科研历程,总的来说它就是一个从现象中发现科学问题,并逐步深入探索的过程。这个过程充满了挑战,又有很多新奇的发现。
从2015年,我开始思考肠道内微生物的进化问题。到2017年,我们开始探索肠道内益生菌的适应性进化。到2019年,我们首先揭示了植物乳杆菌在肠道内适应性进化的现象。到2020年,我们进一步明确了肠道内益生菌进化选择压力的来源。再到2021年,基于实验结果发现了分歧性进化的现象,并对其成因开展探索。直到2022年,确定了基于微生物组定向驯化益生菌的一个可行性。
在接下来的一段时间,我们团队将会更加关注益生菌的驯化问题,就是要解决“So What?”的问题。
也可以跟大家透露一下,我们正在开展的是基于肿瘤微生物组的益生菌定向驯化,我们期待可以驯化出辅助肿瘤治疗的明星菌株。这些内容,我希望以后有机会和大家继续分享。

最后,我要感谢热心肠研究院提供交流的平台,感谢国家自然科学基金、国家重点研发计划、海南省重点研发专项对我们团队的稳定支持,感谢我们团队在这个研究方向的持之以恒探索,还要感谢我们的合作伙伴刘洋彧教授、黄适教授和Rob Knight教授。
感谢聆听!






