

各位热心肠的粉丝,大家好,我是来自云南大学的张志刚。
下面我和大家分享一个非常有趣的故事,就是跨多尺度探讨肠菌演化之谜。

众所周知,地球生命演化的历程给了我们非常重要的启示。
在整个地球诞生的时候,大约在40多亿年前,在我们真核的生物,特别是高等生物出现之前,其实原核的生物已经出现了。这些原核的生物已经帮助我们塑造地球成为能够适合我们的环境,已经塑造了几十亿年的时间。
这个过程当中,除了塑造环境,其实作为共生的伴侣,原核生物对于我们这些地球的生物的形成,以及丰富多样的性状的形成,也起着不可替代的作用。
由此我们认为,微生物其实是非常古老而重要的。

从我们人类认识微生物的历史到现在,才不到400多年。实际上,我们包括动物,已经形成了非常多样的性状。比如说像青藏高原上这些生物,独特的人类还有这些大型的动物,像藏羚羊、金丝猴等等,它们都能够很好地去适应青藏高原独特的环境。

再比如,我们所熟悉的授粉昆虫,它们能够消化蜂蜜当中的果胶,同时也可以为我们丰富多样的开花植物提供授粉服务,帮助这些植物繁衍后代。
我们可以看到,这个图中有非常丰富多样的授粉昆虫。这些性状,其实近来的研究也证实了,它能够传授、消化这些花粉,都是离不开共生的微生物。

上面适应的这些性状与环境有相关,其实我们人类面临的很多的疾病,也是一些复杂的性状。但是这些性状,在我们人类演化历程当中、在衰老的进程当中,是一些不适应的性状,包括一些代谢性疾病、神经性疾病,还有免疫力的疾病,像肥胖,像老年痴呆、帕金森,还有自闭症等等。
近年来的研究发现,这些疾病的发生也与肠菌息息相关。

我们如何去研究这些复杂的性状,肠菌的演化的规律或许能够为我们解析复杂性状形成的机制提供一些新的视角。
其中出发点有两个:我们能否找到一些超级的物种或超级个体,能够具有这些抗逆的性状,然后这些性状又和肠菌密切相关。进而把这些肠菌运用到我们解决复杂疾病的一些问题上来,如何去从肠菌的角度解析复杂性状形成和演化的规律。

我们团队前期以青藏高原上那些天然的物种为例,去展开了研究。
前人研究都认为,青藏高原是一个研究复杂性状的形成和演化的天然的实验室。因为在青藏高原形成的过程当中,欧亚板块不断地碰撞,高原两侧隆升之后,有的动物是从其他大陆进入到青藏高原,有的动物是从青藏高原迁出到其他地方,有一些原住的动物产生了灭绝,有一些也继续留下来。
通过这样周而复始过程,使得青藏高原上这些动物有一个非常漫长的适应青藏高原环境改变的过程,为我们提供了非常好的契机。
这个过程当中,肠道微生物是如何帮助它们适应这个环境的?它们的肠道微生物又是如何演化形成的?我们通过一系列的研究跟大家分享。
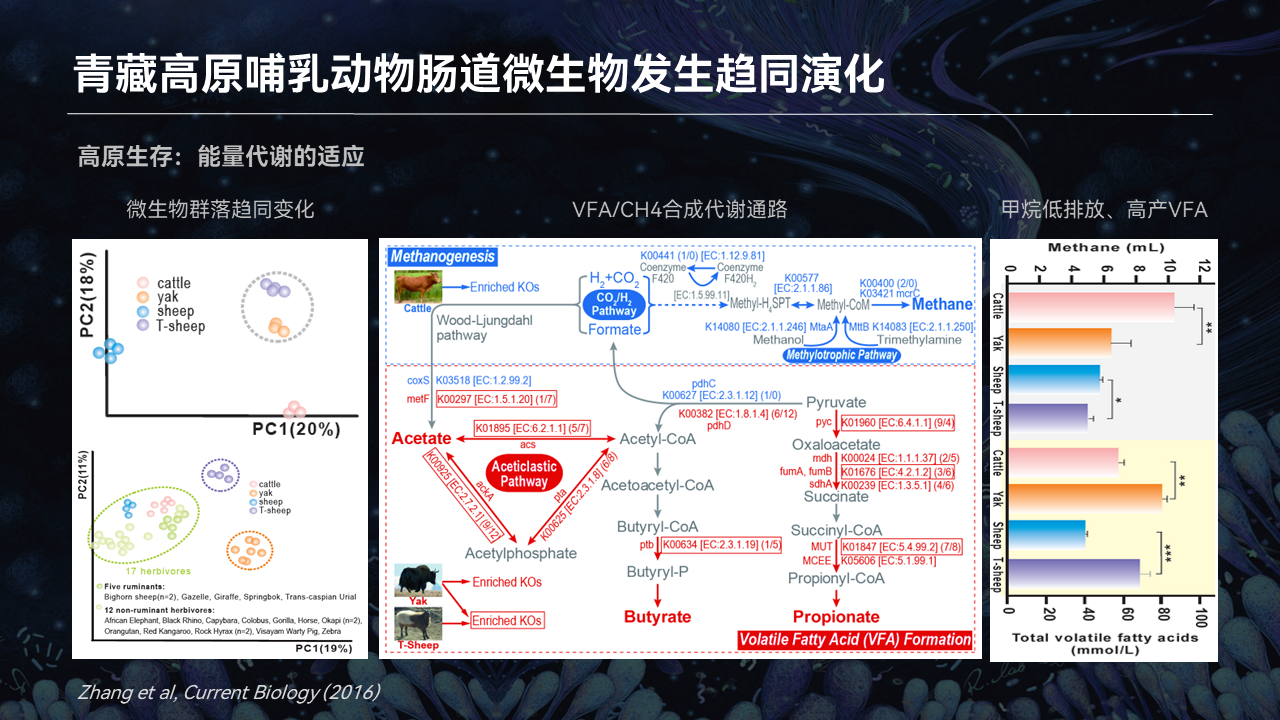
首先我们发现,青藏高原哺乳动物,比如说牦牛和藏绵羊,它们的肠道微生物在结构、功能上一致地区别于低海拔的一些近缘的动物,比如说黄牛,还有一些普通的绵羊。
它这种趋同的演化究竟有什么样的贡献呢?
我们发现,它的肠道微生物能够编码更加丰富的短链脂肪酸产生的这些代谢通路,而在甲烷排放这方面的代谢通路是缺失的。相应的表型上面我们也得到了验证,也就是证明了肠菌能够帮助高原哺乳动物产生更多的能量物质,如短链脂肪酸,同时释放更少的甲烷。
高原哺乳动物,像牦牛还有藏绵羊是一个节能减排的动物。肠道微生物能够帮助它们,从能量的角度来说,很好地适应高海拔环境的一个长期的胁迫。
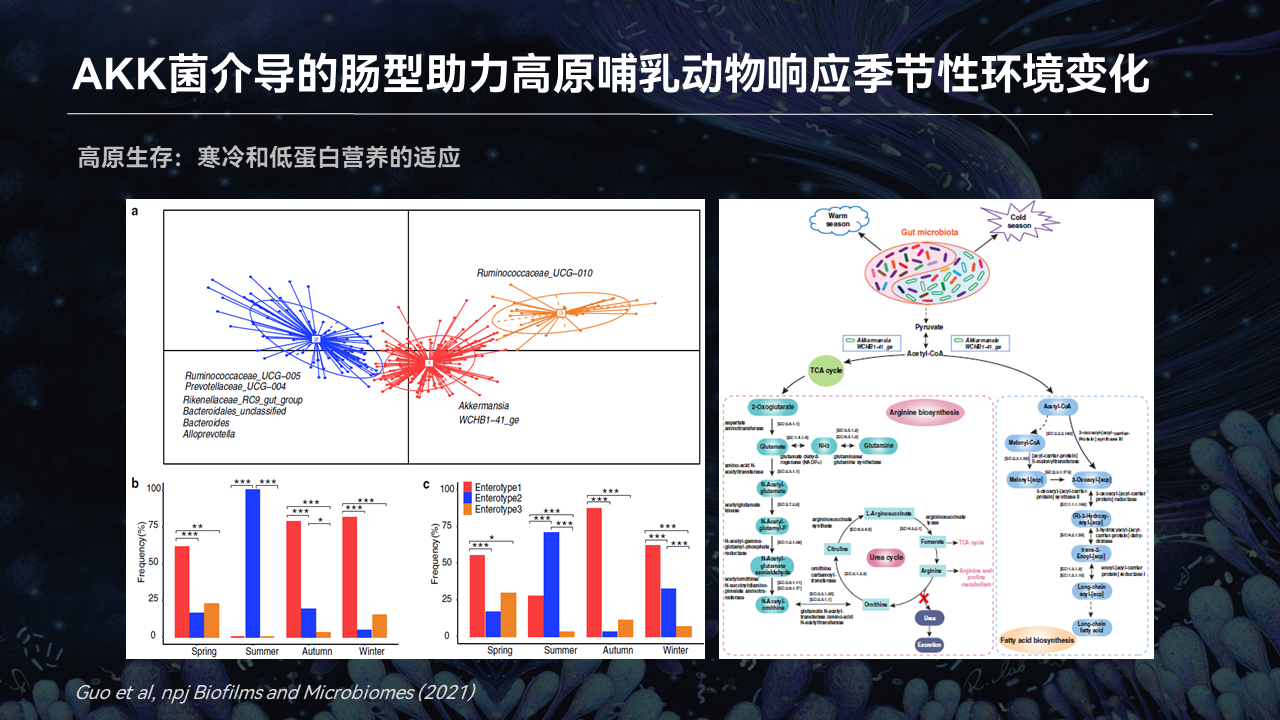
进一步我们发现,在肠道微生物当中,如何能够帮助高原动物能够适应季节性的一些食物的改变,还有冷暖的改变。我们在牦牛当中发现了什么?
我们熟悉的阿克曼菌(AKK菌),它就介导了一个肠型。这个肠型在冷季,除了热季就是6月份到8月份,剩下都是冷季,一致地都要高于暖季,也就是图中的红色的柱子。
这些菌都是在冷季介导的肠型非常多,阿克曼菌介导的肠型究竟有什么价值?最后我们解析阿克曼菌的整个代谢通路,有些可以合成短链脂肪酸、合成高级脂肪酸,它有助于在冷季能量的积累。同时它在鸟氨酸循环当中产生尿素这一步的基因是缺失的,反而它就可以把碳水化合物转化成氨基酸,而不以尿素的形式去丢失。
这对于牦牛或者高原哺乳动物来说,有什么价值呢?实际上在青藏高原冷季的时候,与暖季比起来,食物当中的蛋白含量下降了很多。在冷季的时候,它需要更多的能量来去适应。
阿克曼菌介导的这个肠型正好能够帮助它来储存能量,同时满足食物当中低氮或者是低蛋白营养的一个胁迫。
这个研究进一步证明了,肠菌是可以帮助高原哺乳动物响应季节性的对能量适应的要求,还有低氮、营养胁迫的需求。
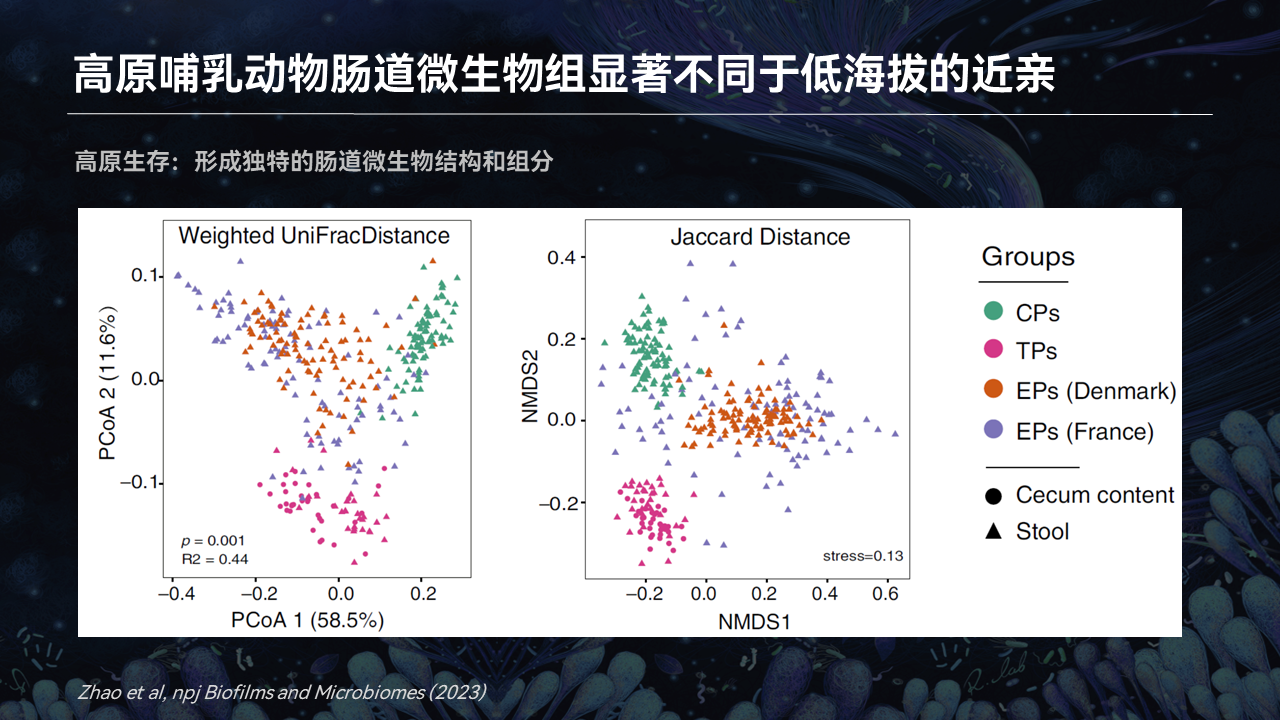
除了反刍动物,别的单胃动物它是不是也是有类似这样的情况呢?
我们发表的研究也对藏猪(TP)和其他的我们中国圈养的家猪(CP)以及欧洲的家猪(EP)进行了比较以后,我们发现藏猪的肠道微生物,它都是不同于欧洲家猪和中国家猪。
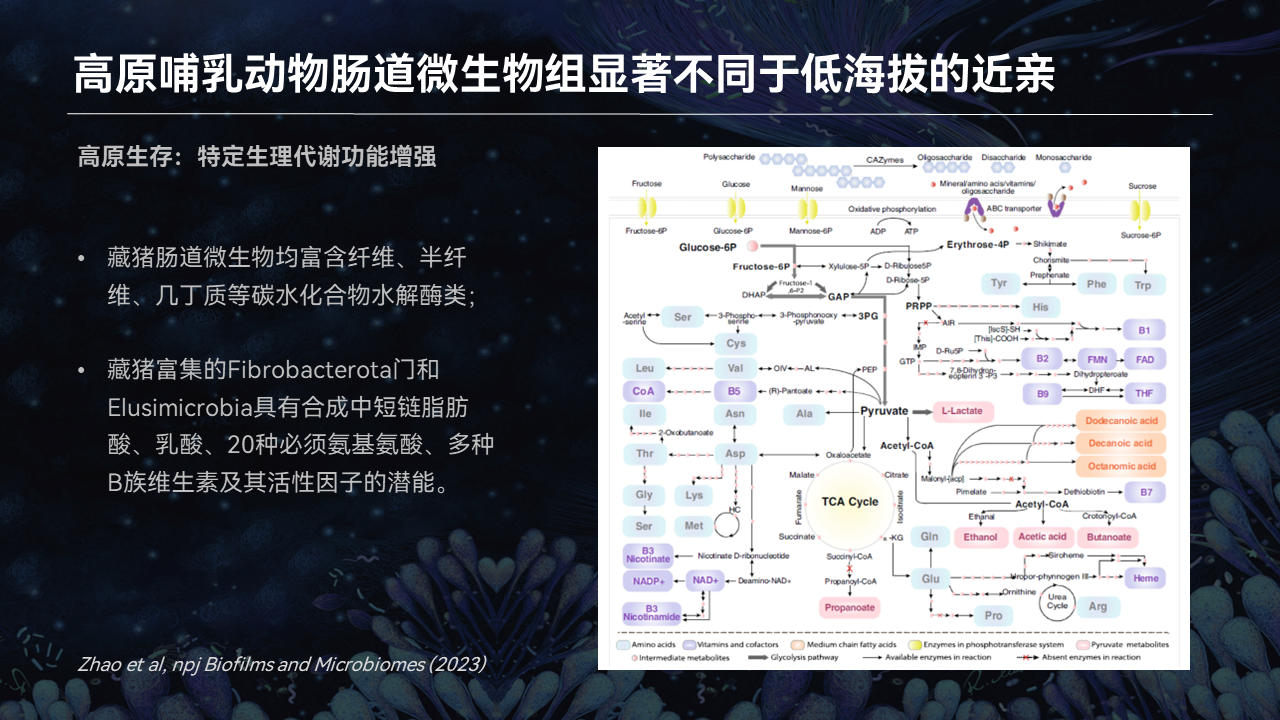
同时我们在深度解析藏猪的肠道微生物的时候,我们发现它富集的这些肠菌,能够编码丰富的纤维素、半纤维素、几丁质等等复杂多糖降解的水解酶。
同时它富集的这两个细菌门,都能够合成各种短链脂肪酸、乳酸以及必需的20多种氨基酸,还有一些丰富的B族维生素等等。
这些新陈代谢的能力,能够帮助单胃动物藏猪很好地去适应高海拔能量的需求和对低氧胁迫的一个需求。
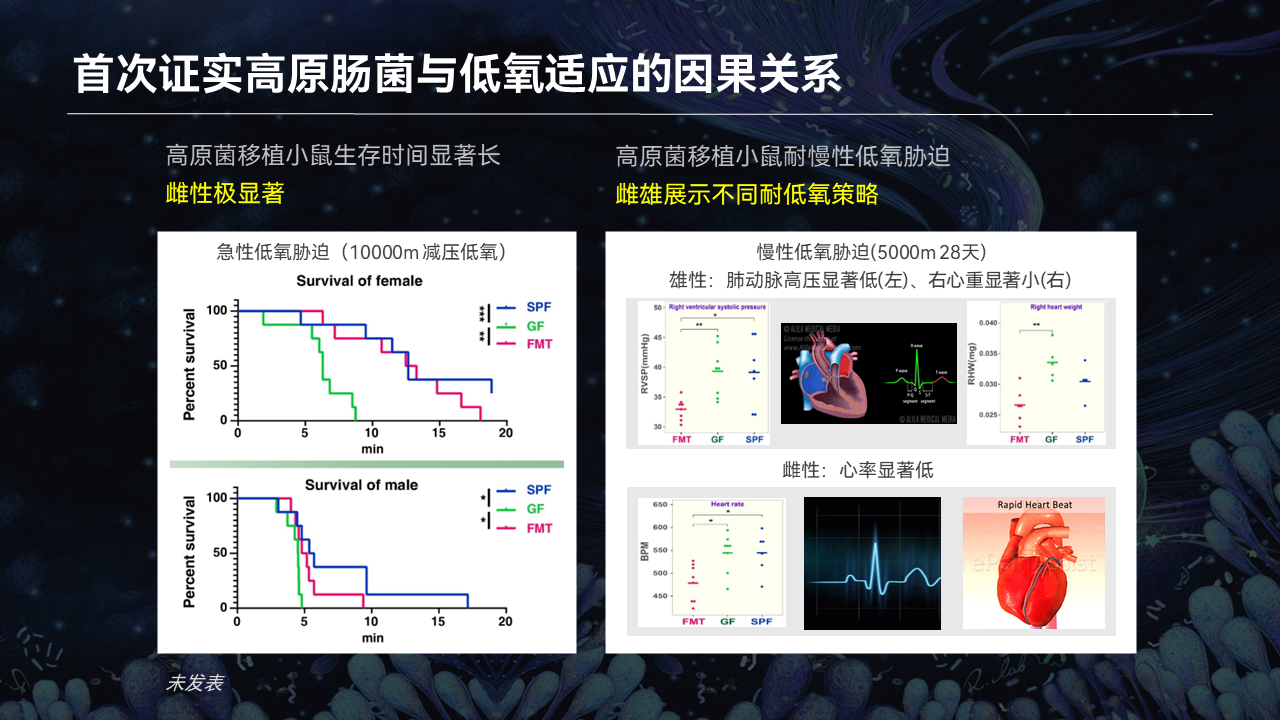
那么人群上面究竟怎么回事?在高原环境当中一个重要的普遍的环境胁迫因子就是低氧,肠菌与低氧有什么关系?
我们首次也证实了,通过FMT(粪菌移植)把高原肠菌移植到无菌小鼠,做了急性低氧胁迫和慢性低氧胁迫,我们证实了高原肠菌与低氧适应之间的因果关系。
图中左边,我们是做了急性低氧,模拟1万米海拔的急性低氧。我们发现移植我们高原肠菌生存率是显著的提升,特别在雌性当中表现更为明显。
在慢性模拟5000米、1个月的持续胁迫下面,我们看到高原肠菌都表现出抗低氧胁迫的性状,当然也是雌雄有别的。
这些所谓抗低氧的性状是什么呢?比如说抗肺动脉高压、抗心肌肥厚、抗心律异常。这些信息为我们提供了一个重要的参考,就说特定的高原肠菌与高原低氧胁迫是密切相关的。

这些菌究竟是什么呢?
我们进一步比较了3个海拔的藏族人群的肠道微生物。最右边的是4000米海拔以上的,还有3500米的,还有1800米左右的。
我们发现,毛螺菌类的成员以及少数的柔嫩菌属的成员,它与藏族人群高原适应是密切相关的。这个是没有发表的成果。

进一步的话,如果我们低海拔的人群上去高原以后,有哪些菌能够特异地与我们响应高原习服有关呢?
我们还是发现了,肠道的毛螺菌是与平原人群应激快速的高原低氧胁迫是密切相关的,其中这里面典型的代表就是布劳特菌。
这个试验,我们比较了重庆的军人上了高原以后,特别是拉萨,一个月两个月以后我们去收集采样,以及回到重庆以后再去收集样。通过这样系列的一个比较,发现了这样的规律。
由此可见,毛螺菌科的成员是与我们高原低氧胁迫密切相关的。目前确确实实我们也从藏族人群当中分离到了系列的毛螺菌的菌株,正在开展单菌水平的因果的论证以及机制的解析。
我们可以看到,高原哺乳动物的肠道微生物,它能够很好地贡献于高原哺乳动物适应高原的极端环境,包括低氧、冷以及能量的适应。

它的肠道微生物是如何演化的?符合什么样的规律?
为了很好地去回答这个问题,我们去收集了7000多份在高原人群和动物的样本的资源库。
为了更好地去了解这些动物生存的环境,我们也拍摄了首部科考纪录片,叫《卓乃湖》,来再现这些多样化的丰富多样的动物,它生存的生境以及气候特征。

在样本资源基础上,我们就构建了一个高原动物的微生物资源库。这里边包括2000多个宏基因组的数据,数据量也达到了50多TB以上。特别是我们已经获取了2万多个物种水平的中高质量的参考基因组,包括了细菌和古菌,新种是代表了差不多都是在90%以上。
这个为我们解析上面的问题提供了非常好的一个保障。
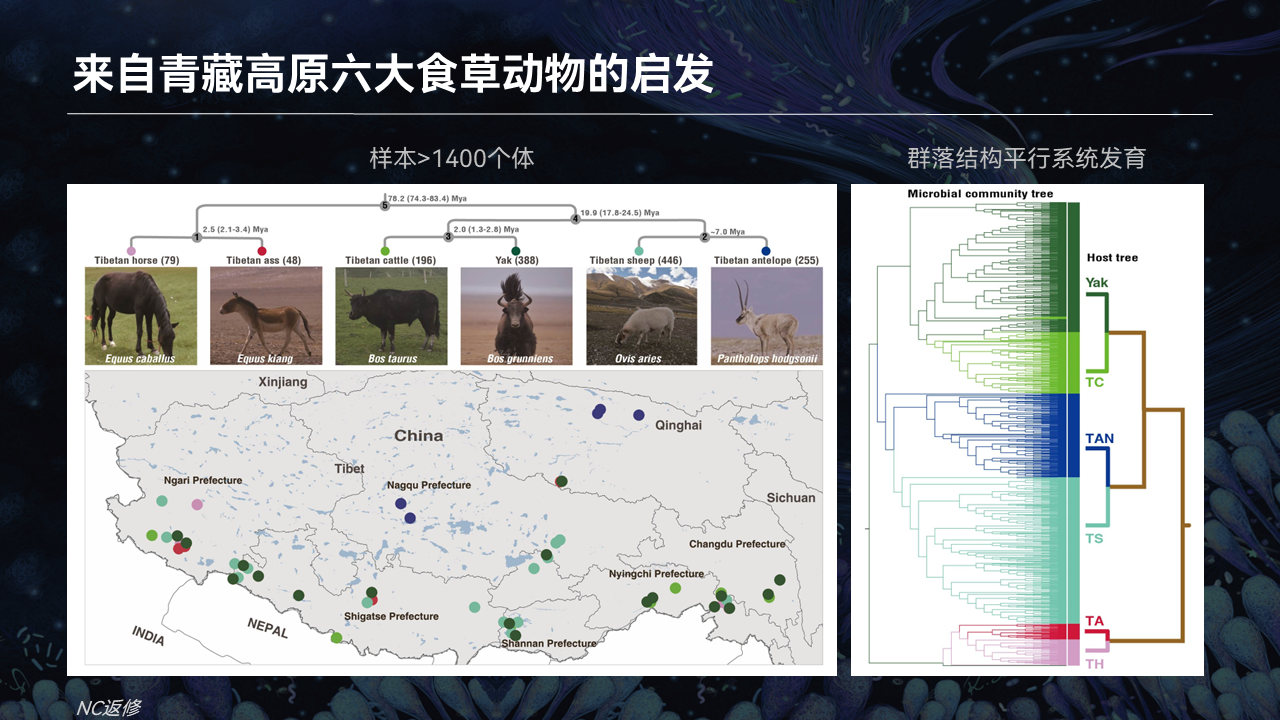
基于这些数据,我们利用了6个大型的食草动物,近亲的食草动物,包括奇蹄目的藏马和藏野驴,偶蹄目的牦牛、藏黄牛、藏羚羊和藏绵羊这6个物种,来去通过大的数据的深度测序,我们调研它整个演化的规律。
首先我们看一下,它的肠道微生物的群落结构。右图展示的一个群落结构,它正好是平行于动物宿主的系统发育关系。
也就是说在奇偶蹄目这样的一个进化时间尺度上面,宿主的系统发育或者宿主遗传的这种分化,它是对肠群落结构的形成起到了非常大的一个约束的作用。
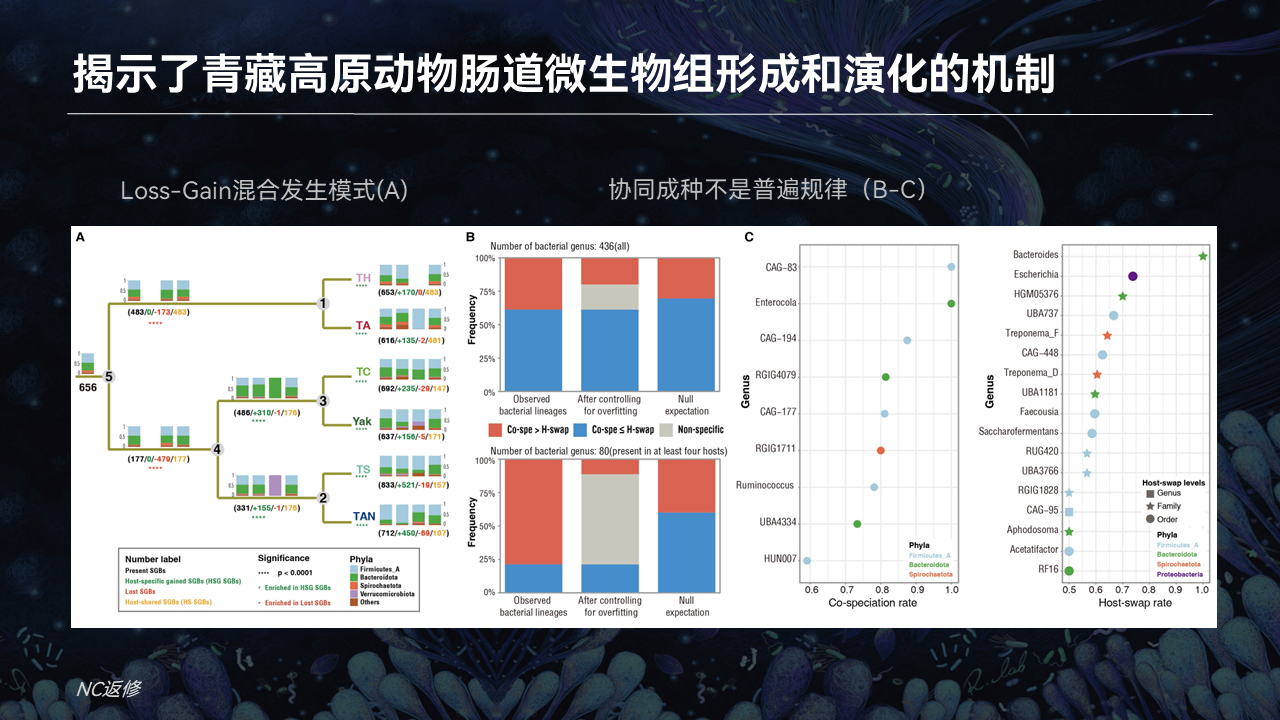
在肠道微生物物种形成的过程,它究竟是一个什么样的规律?
首先我们看左边的A图,它展示出了一个与预测的共同祖先相比,它是通过早期的丢失和近期的重新获得形成的最终稳定的肠道微生物群落结构。
在肠道微生物物种形成过程当中,不同以往理论的认知,这些肠菌的形成至少协同成种,也叫Co-speciation(共同物种形成),或者Co-phylogeny(共同系统发育)。
平行于系统发育关系这些菌,这样类型的事件其实并不是最普遍的。也就是其他的事件,比如说是跨宿主的水平转移这种事件,以及丢失或者是获得这种事件,它其实是占据非常非常重要的地位的。
这是从一个小尺度展示了哺乳动物肠道微生物形成和演化的规律。

我们更希望看到,在更大的一个尺度上面,肠菌形成的一个普遍规律是什么样子。因此,我们就做了一个全球最大规模授粉昆虫肠道微生物的分析。
这里面包括了36个物种,属于4个目,基本涵盖了主要的授粉昆虫的一个类群,涉及到鳞翅目、半翅目、膜翅目这些昆虫。
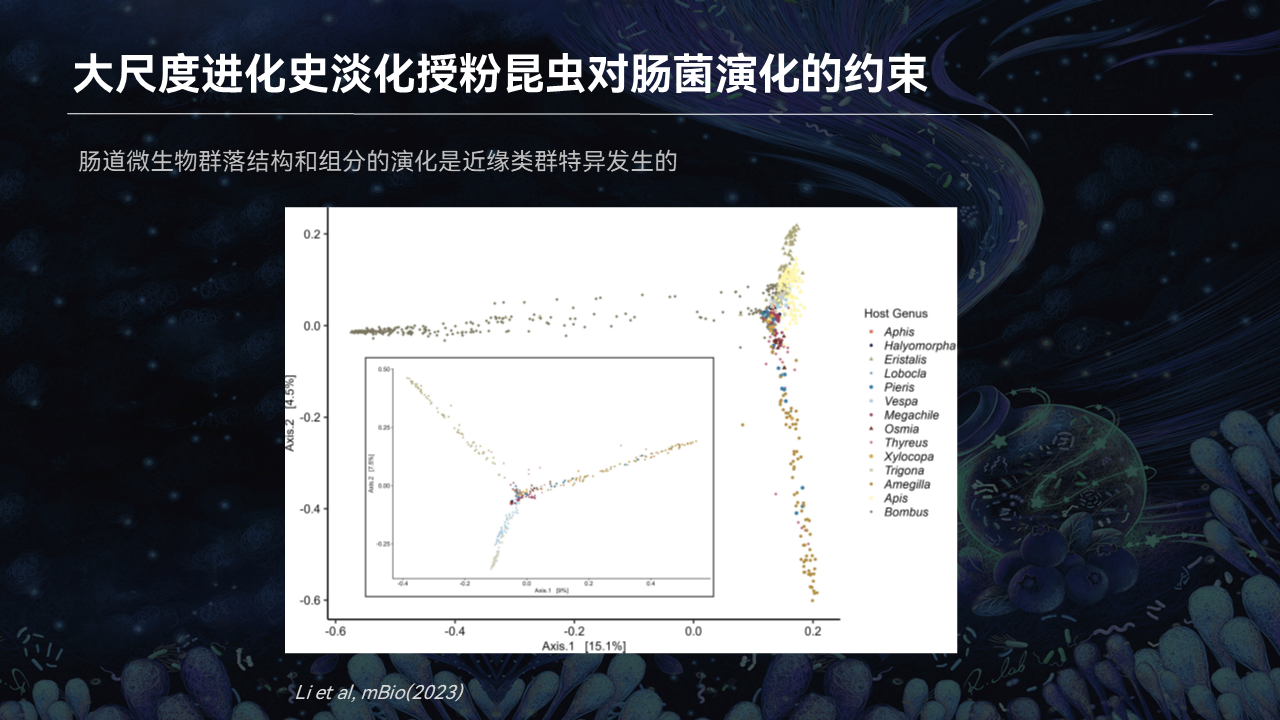
我们在解析了它的肠道微生物结构基础上,去看了一下它整个肠菌群落结构演化的规律。
结果我们很吃惊地发现,在非常大尺度的宿主系统分化基础上,群落结构的形成和演化已经脱离了宿主遗传的一个曲线,或者脱离了宿主系统发育的限制,更多地可能受到了环境、食物等等多方面因素的影响。
那么这个也是不同于以往的一个基础的认知。

我们不管从小的尺度和大的尺度,我们都看到了肠道微生物能够很好地帮助动物宿主适应环境。那么它的形成规律是一个什么样子的,从群落结构以及到肠道微生物组形成的一个机制。
接下来,我们非常好奇一个问题:不管什么样的动物,我们的肠道微生物究竟是如何起源演化的?这是一个非常值得思考的问题,我们如何站在一个全球的尺度来去认识这个问题。
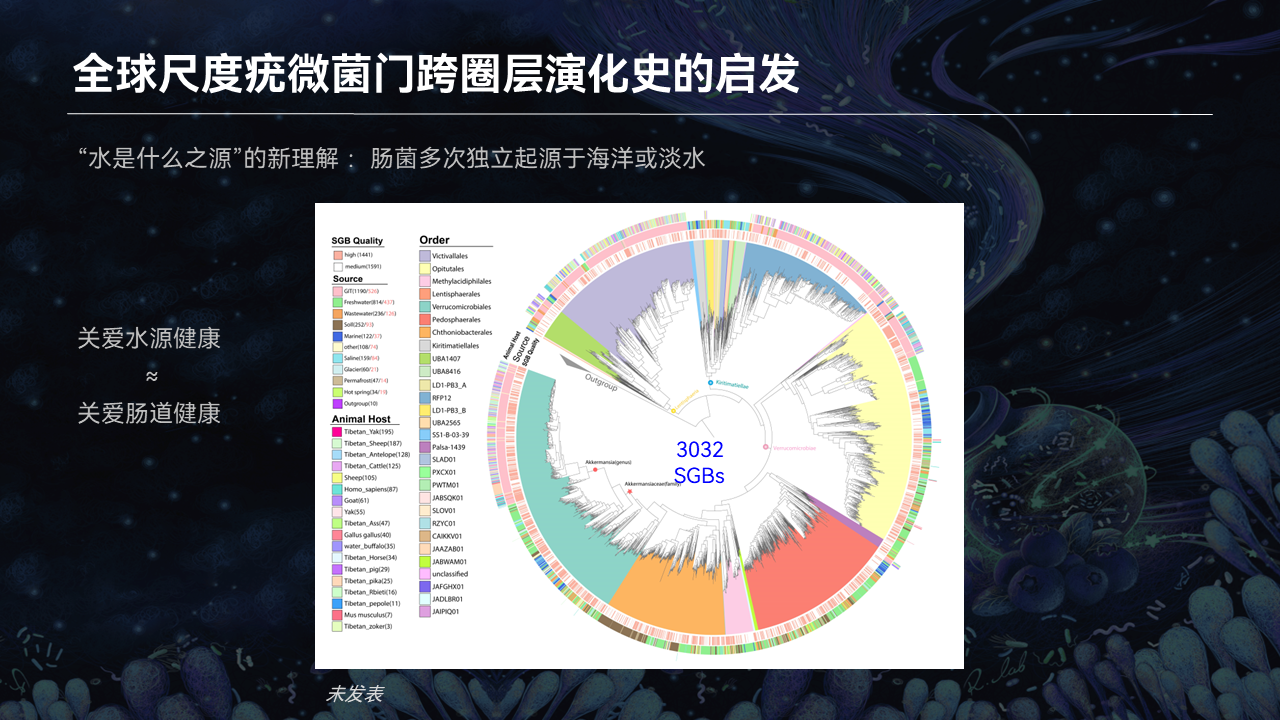
我们正好是利用我们所熟悉的肠道疣微菌门这样一个系统的类群,去在全球尺度、不同圈层来探索肠菌的演化的一个规律。
从图中我们可以看得到,粉色的部分全是我们肠菌,在早期演化的过程当中,最近亲的要么就是海水,要么就是淡水上面的这些疣微菌门。
由此可见我们肠菌的话,可能的早期来源是海水。我们可以看到,演化树的右上部分还有右边这个部分,这是相对说早期一点的。我们可以发现,肠菌它是多次独立地由海洋近亲的这些菌去进化出来的。
我们绕着顺时针走的话,我们看到有意思的是有一些类群是肠菌,它可能是被排放到这个环境当中,在环境当中又形成独立的一支,比如说黄色的这一块,右下角橙色和黄色这块是属于土壤的菌。然后我们再往左边看,我们发现,肠菌它又是独立地演化成一个类群,它的近亲又是来自于淡水。
其实我们可以得到第一个初步的结论,我们的肠菌的来源,它是多次独立演化出来的,而不是一次从一个祖先的状态过来的。
那么最重要的环境来源是什么呢?早期的这些形成的类群,它是属于海洋。近期的这些类群的话,淡水是非常非常重要的。
大家所熟悉认知的一个理念就是“水是生命之源”。其实我们当前的研究,也进一步能够提出一种新的认知,对于我们肠菌的来源其实水体也是非常重要的。
有鉴于此,我们就提出这样一个理念,其实关注全球的水资源的健康和水体的健康,也就是关注我们的肠道的健康,最终也就是关注我们自己的健康。
这个问题,我们仅仅是从疣微菌门这个分类,从全球尺度上来去探索它是如何起源演化的。在其他方面、其他的一些菌,它是什么样的规律,我们后续还会进行一个继续的探索。

前面我们给大家分享的,是在一个独特生境当中形成一个非常复杂多样的复杂性状,菌群有什么贡献,它是怎么去演化形成,它是怎么去起源的,提供了非常好的一些系统的研究思路。
实际上,从肠菌的角度来探索、寻找、解析复杂性状形成之谜,其实也是任重而道远的。
不管我们去利用这个群体水平找超级个体,还是跨物种比较找超级物种来说,都是摆在我们众多人面前的一个非常巨大的挑战。

最后我们这些项目,致谢支持我们的国家基金委、科技部等等,以及跟我们合作的昆明动物研究所、华大基因等等这些单位,也感谢云南大学云南生物资源保护与国家利用重点实验室的平台。

最后,也欢迎我们肠菌的粉丝来加入我们的实验室,让我们一起去探索非常有趣的生物多样性的世界!






